Нефрологія для сімейного лікаря - О.Й. Бакалюк 2003
Структурно-функціональна організація сечовидільної системи
До складу сечовидільної системи входять дві нирки, два сечоводи, Сечовий міхур і уретра.
Нирки - парний орган специфічної форми. В них розрізняють передню і задню поверхні, медіальний і латеральний краї, верхній і нижній полюси. Посередині медіального краю знаходяться ворота нирки (hilus renalis), які ведуть у порожнину під назвою sinus renalis. Через ворота в нирку проникають та виходять із неї кровоносні, Лімфатичні судини, нерви.
Нирки розміщені на рівні ХІІ грудного, І та ІІ поперекових хребців по обидві сторони від хребта, за очеревиною й оточені товстим жировим шаром. Права Нирка фіксується дещо нижче від лівої, а верхні полюси нирок ближчі один до одного, ніж нижні (рис. 1).
Обидві нирки важать близько 300 г, причому ліва нирка важча, ніж права. Відношення маси обох нирок до загальної маси тіла складає 1 : 240. Нирки обмежено рухомі у вертикальному напрямку і у своєму нормальному положенні утримуються сполучнотканинною (ниркова фасція) та жировою капсулами, які їх оточуюють, тонусом м’язів передньої черевної стінки. Роль судин у процесі фіксації нирок відносно невелика.
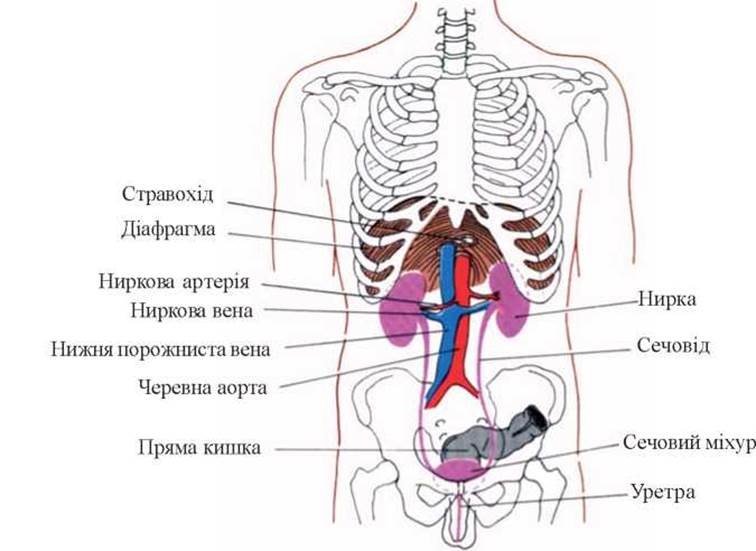
Рис.1. Топографія нирок.
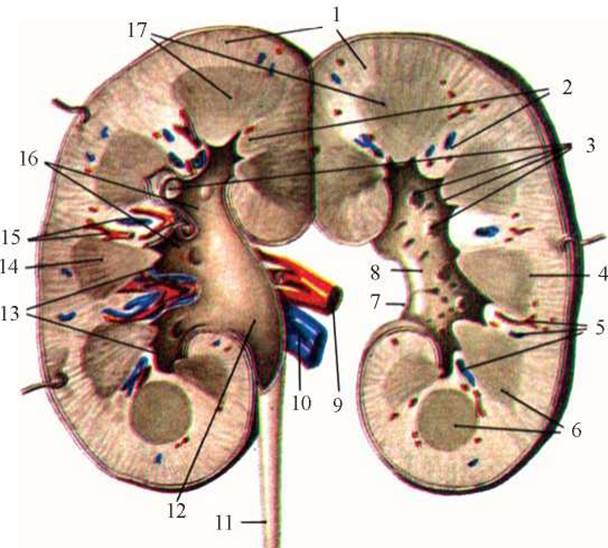
Рис. 2. Фронтальний розріз правої нирки (схема): 1 - кіркова речовина, 2 - ниркові стовпи, 3 - ниркові сосочки, 4 - основа піраміди, 5,15 - Кровоносні судини, 6,14 - ниркові піраміди, 7 - ниркові ворота, 8 - ниркова пазуха, 9 - ниркова артерія, 10 - ниркова вена, 11 - Сечовід, 12 - ниркова миска, 13 - малі ниркові чашечки, 16 - великі ниркові чашечки, 17 - мозкова речовина нирок.
При поздовжньому розрізі нирки від латерального до медіального краю можна побачити, що її ворота ведуть у центрально розміщену порожнину - ниркову пазуху. У цій порожнині розміщуються значна частина ниркової миски, великі та малі чашечки, Кровоносні та лімфатичні судини, нерви; там же видно верхівки ниркових пірамід у вигляді сосочків - papillae renales. На поверхні сосочка знаходяться невеликі отвори - foramina papillaria, через які витікає сеча (рис. 2). Мозкова частина (25-30 % від усієї маси нирок) відрізняється від кіркової тим, що вона забарвлена в менш насичений блідочервоний колір.
Морфологія і функція нефрона детально описані (Г. Маждраков и соавт., 1980; Ю.В. Наточин 1994; В.В. Серов 1995; A. Bohle et al. 1989), тому ми обмежимося викладом лише основних даних, необхідних для розуміння структури і функції нирок.
Структурною одиницею нирки є нефрон. Загальна кількість нефронів в обох нирках сягає 2-2,5 млн, 4/5 їх кількості розміщені у кірковій речовині, 1/5 - на межі кіркового і мозкового шарів. У структуру нефрона входять судинний клубочок, його капсула і нирковий канадець. Схематична будова нефрона показана на рисунку 3.
У нирках людини знаходять два різновиди нефронів - кортикальні (поверхневі) і юкстагломерулярні (глибокі). Юкстагломерулярні нефрони містять у своєму складі петлю Генле, яка доходить до ниркового сосочка. Існує думка, що наявність двох типів нефронів пов’язана з необхідністю надзвичайно тонкої регуляції вмісту в крові натрію як основи електролітного гомеостазу організму (W. Cupples et аІ., Д996).
До складу судинного клубочка входять близько 50 капілярних петель (рис. 4).
Вигляд ниркового клубочка під електронним мікроскопом подано на рисунку 5.
Стінка капіляра є клубочковим фільтром, який складається з епітелію, ендотелію і розміщеної між ними базальної мембрани (БМ).

Рис. 3. Схематична будова нефрона.
Ендотелій капілярів клубочків містить пори розміром 100-150 нм - основний шлях для ультрафільтрації.
БМ капілярів (її товщина - 250-400 нм) складається із трьох шарів - lamina densa, lamina rara externa, lamina rara interna. Власне базальною мембраною є середній шар - lamina densa. До складу БМ входять високоспецифічні білки (колаген IV і V типів, ламінін, ентактин, нефритогенозид, фібронектин, негативно заряджені гепарансульфатпротеоглікани).
Важливе значення для забезпечення функції БМ надають колагену IV типу.
У БМ міститься 6 різних варіантів ланцюгів колагену цього типу, які позначаються відповідно від а1 до а6. Кожен ланцюг кодується своїм Геном, і ці гени розміщені парами на трьох різних хромосомах. Власне з генетичними порушеннями структури двох із шести ланцюгів колагену IV типу пов’язані клінічні синдроми Альпорта і Гудпасчера (див. нижче). БМ теж містить пори (середній розмір - 2,9 нм), розмір яких може змінюватися залежно від величини гідростатичного тиску в капілярах клубочка. БМ не є суцільною, вона переривається у тих місцях, де розміщені ядра ендотеліоцитів, і у цих прошарках у БМ «вставлені» мезангіоцити. Білкові складові БМ формують і відповідний електростатичний заряд, зміна величини якого (зменшення) має суттєве значення для виникнення протеїнурії (ПУ).
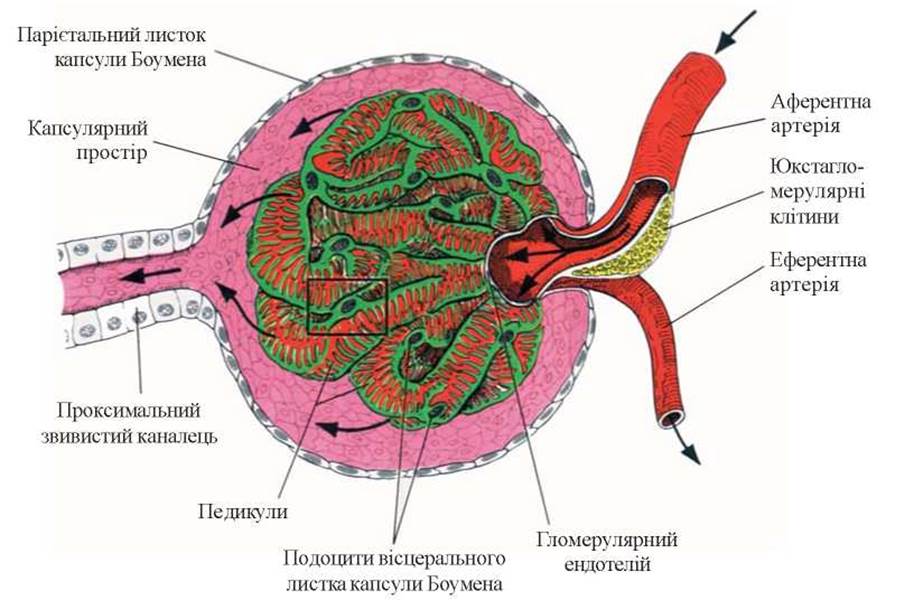
Рис.Э. Схематична будова клубочка.
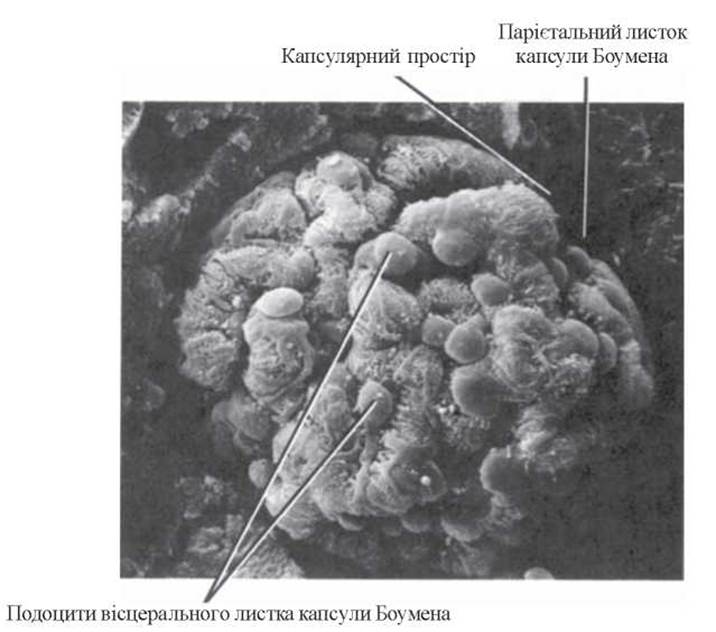
Рис.3. Ширковий клубочок. Скануюча електронна мікроскопія, х 1230 (R.G. Kessel et al., 1979).
Епітеліальні клітини (подоцити) нагадують подушку з відростками (або спрута), яка покриває капіляр зовні (рис. 6).
Малі відростки подоцитів (педикули) відходять перпендикулярно від великих і тісно переплітаються між собою, утворюючи разом із дрібними фібрилярними структурами так звану щілинну діафрагму (pecular diaphragma). Маючи філаментарну будову, щілинна діафрагма утворює, таким чином, своєрідну систему пор (діаметром 5-12 нм), якій теж відводять надзвичайно важливу роль у процесі гломерулярної фільтрації.
Вигляд подоцитів під електронним мікроскопом наведено на рисунку 7.
Капілярні петлі клубочка підвішені на зразок брижі кишечника в ділянці полюса клубочка мезангієм - сполучнотканинною структурою, до складу якої входять клітини-мезангіоцити і мезангіальний матрикс. Дослідження останніх років дозволили визначити важливу роль мезангія в забезпеченні функції нефрона в нормі і в її порушенні при патології.
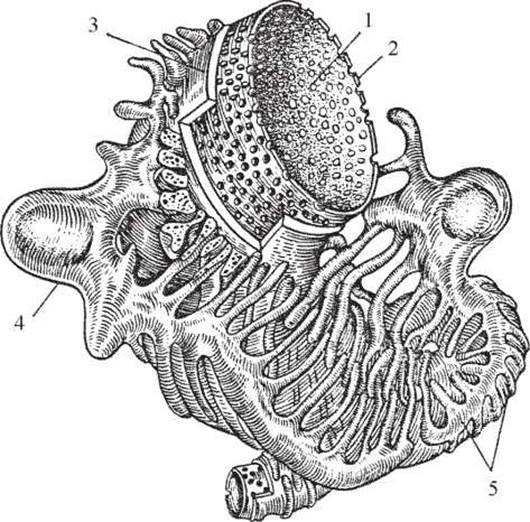
Рис.6. Гломерулярный фільтр. Схема будови стінки капіляра (за М.Я. Ратнер и соавт., 1987): 1 - просвіт капіляра; 2 - ендотелій; 3 - базальна мембрана; 4 - подоцит; 5 - мазі відростки подоцитів (педикули).
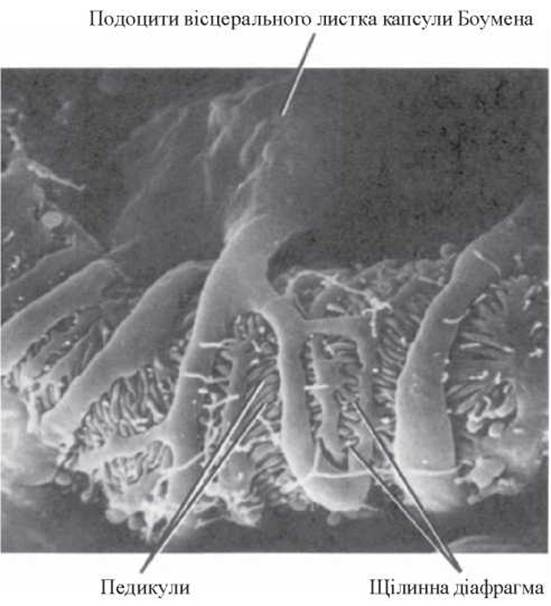
Рис.7. Подоцит. Скануюча електронна мікроскопія, х 7800 (R.G. Kessel et al., 1979).
Виділяють 2 групи мезангіальних клітин - власне гломерулярні (контрактильні) і транзиторні (резиденті) мононуклеарні лейкоцити.
Контрактильні мезангіальні клітини, які складають основну масу клітин мезангія, мають багато спільних властивостей із гладеньком’язовими клітинами судинних стінок і відіграють важливу роль у регуляції капілярного кровообігу та процесу ультрафільтрації. Ці клітини містять актоміозин, рецептори до ангіотензину-ІІ (АТ-ІІ). За певних умов вони здатні виконувати функції амілоїдокластів, продукувати ренін, скорочуватися при дії на них вазопресину, паратгормону, простагландинів (ПГ) І2 та Е2, гістаміну, що призводить до зменшення загальної фільтраційної поверхні клубочка (J.I. Kreisberg et al., 1995). Одночасно ці клітини синтезують колаген І, ІІІ, ІV, V типів, фібронектин, глікозаміноглікани, секретують нейтральні протеази, зокрема колагеназу, та специфічні деградуючі компоненти матриксу. Таким чином, контрактильні мезангіальні клітини забезпечують поновлення мезангіального матриксу та його фізіологічний апоптоз.
Транзиторні мезангіальні клітини складають 3-7 % усієї популяції, in vivo та in vitro вони фагоцитують циркулюючі імунні комплекси (ЦІК), частинки латексу, бактерії, агрегований гаммаглобулін.
Гломерулярний мезангій має власну, відносно незалежну від капілярного кровобігу, гемоциркуляцію (A.F. Michael et al., 1990). Це дозволяє стверджувати, що навіть невелике число мезангіальних мононуклеарних фагоцитів, виділяючи БІОЛОГІЧНО АКТИВНІ РЕЧОВИНИ, продукція яких переважно характерна для циркулюючих тканинних моноцитів-макрофагів, може впливати на стан усієї мезангіальної ділянки. Таким чином, присутність у нормальних клубочках незначної кількості мезангіальних клітин створює умови для локальної реалізації імунних реакцій, результатом яких є потовщення та склероз стінки капіляра або повна облітерація його просвіту.
Основними компонентами мезангіального матриксу теж є колаген ІV і V типів, ламінін, фібронектин. У функціональному відношенні матрикс виконує роль каналу («сміттєпроводу») для видалення продуктів розпаду білкових молекул, мікроорганізмів або їх фрагментів.
Капсула клубочка представлена БМ і епітелієм. БМ капсули є початком БМ стінки канальців, а епітелій капсули за своєю функцією нагадує епітелій головного відділу канальців. Епітелій капсули теж має здатність до скорочення, а отже, до зміни об’єму капсули.
Канальцеву частину нефрона поділяють на головний проксимальний відділ, петлю Генле, дистальний відділ, збиральні трубки. Ендотелій кожного із цих відділів характеризується специфічною ультраструктурою, яка сприяє максимальному виконанню ними своїх функцій - реабсорбції, секреції тощо.
Вигляд канальців під електронним мікроскопом подано на рисунку 8.
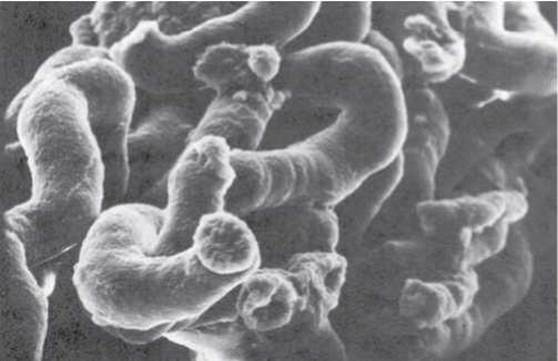
Рис. 8. Ниркові канальці. Скануюча електронна мікроскопія, X 500 (за Courtesy of Fisher Scientific Company and S.T.E.M. Laboratories, Inc., Copyright, 1975).
У місці контакту звивистої частини дистального канальця і полюса судинного клубочка епітелій цього відділу змінює свою структуру. За пропозицією W. Zimmerman (1933), ця частина канальця отримала назву macula densa. Клітини цього відділу надзвичайно тісно контактують зі структурними елементами юкстагломерулярного апарату (рис. 9).
До складу юкстагломерулярного апарату входять:
- епітеліоїдні клітини стінок приносної та виносної артерій клубочка;
- клітини Гурмагтіга (lacis-клітини), які розміщуються у просторі між приносною і виносною артеріями клубочка;
- мезангіальні клітини клубочка.
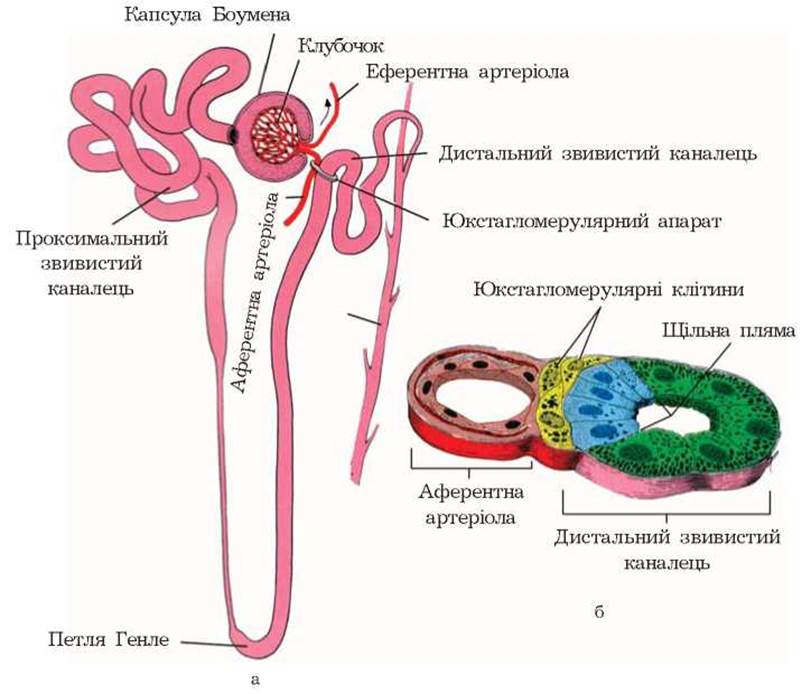
Рис. 9. Схема будови юкстагломерулярного апарату нирок: а - розміщення; б - поперечний розріз.
Тісний контакт macula densa з іншими складовими юкстагломерулярного апарату забезпечує вплив хімічного складу сечі в просвіті дистального канальця на клубочковий кровобіг і внутрішньоклубочковий тиск і, навпаки, реалізує гормональні впливи юкстагломерулярного апарату на функцію клітин macula densa.
На рівні окремого нефрона ця ниркова структура виступає як локальний регуляторний механізм. Завдяки гломерулярно-тубулярному шунту вона постійно ніби порівнює величину клубочкової фільтрації (КФ) і склад плазми, яка фільтрується в клубочку, зі складом канальцевої порції сечі. За принципом роботи юкстагломерулярний апарат нагадує водомірне скло парового котла з тією відмінністю, що індикатором є не рівень рідини, а кількість іонів натрію у плазмі і в сечі. При невідповідності порівнюваних параметрів юкстагломерулярний апарат може автоматично, шляхом вивільнення реніну та локального синтезу АТ-ІІ, зменшити просвіт привідної Артерії клубочка, змінивши тим самим фільтраційне навантаження і привівши його у відповідність із реабсорбційними можливостями канальців.
Збиральні трубки забезпечують пасивний транспорт води, секрецію іонів водню (Н+).
До ендокринної системи нирок належать система ПГ, калікреїн-кінінова система, а також апудоцити. ПГ продукуються інтерстиціальними клітинами мозкової речовини і нефроцитами збиральних трубок, складові калікреїн-кінінової системи - нефроцитами дистальних відділів канальців. Ниркові апудоцити (онкоцити і М-клітини) є складовою клітинної організації з високим ступенем активності - так званої APUD-системи. Остання локалізована в різних органах, які мають або не мають гормональну активність (Епіфіз, Гіпофіз, щитоподібна, паращитоподібні, Надниркові залози, Печінка, Підшлункова залоза, Стравохід, Трахея, Бронхи, смакові сосочки язика тощо). Ниркові апудоцити продукують біогенні аміни, які виконують функції як гормону, так і медіаторного нейраміну (дофамін, меланотонін, серотонін).
Виконання нирками життєво важливих функцій є неможливим без їх адекватного кровопостачання. Нирки належать до органів, які найкраще постачаються Кров’ю, - за 1 хвилину через нирки проходить до 25 % хвилинного об’єму крові (з коливаннями від 12 % до 30 %), що складає в середньому 1200 мл/хв для людини з масою тіла 70 кг. Величина кровотоку в нирках за інтенсивністю перевищує м’язовий кровобіг у спокої у 100 і більше разів. Особливістю ниркового кровобігу є також його здатність до авторегуляції і змін КФ у досить широкому діапазоні практично незалежно від рівня системного артеріального тиску (АТ).
У нирках існують два кола кровобігу - кортикальне і юкстамедулярне (рис. 10).
У фізіологічних умовах 85-95 % крові проходить по кортикальному (великому) і 3-10 % - по юкстамедулярному (малому) колах. Ці два кола відокремлені одне від одного за допомогою vas efferens, які чинять значну перешкоду току крові. В результаті цього в клубочковому капілярному руслі зберігається досить високий тиск (у vas efferens близько 70 мм рт.ст., ниркових капілярах - 45-60 мм рт.ст., нирковій капсулі і звивистих канальцях І порядку - 15 мм рт.ст., петлі Генле - 10 мм рт.ст., інтерстиціальній рідині - 10 мм рт.ст.), а у перитубулярному — низький (у дистальних канальцях - 6 мм рт.ст., збиральних трубках - 2-0 мм рт.ст., перитубулярних капілярах - 13 мм рт.ст., вивідній нирковій вені - 45 мм рт.ст.) (рис. 11).
Низький тиск у перитубулярному капілярному руслі і сповільнення току крові в ньому створюють сприятливі умови для процесу реабсорбції різних речовин.
Невелика (23 %) кількість крові, яка доходить до нирки, може проходити через артеріовенозні анастомози (шунти). Її об’єм може значно зрости за певних умов (нейрорефлекторні і гормональні зміни) і спричинити скид крові безпосередньо в піраміди, обминаючи кірковий шар, що може призвести до розвитку його ішемії та некрозу.
Лімфатична система нирок функціонально підпорядкована реабсорбційній функції канальців. Більш тісний контакт лімфатична система має з артеріальною системою, менш тісний - із венозною. Лімфатичні судини відсутні в гломерулах і пірамідах, і в цілому анатомічно лімфатична система нирок певною мірою відповідає конструкції їх кровоносної системи. Початкові елементи лімфатичних капілярів у вигляді сіточки оточують капсулу клубочків, канальці.
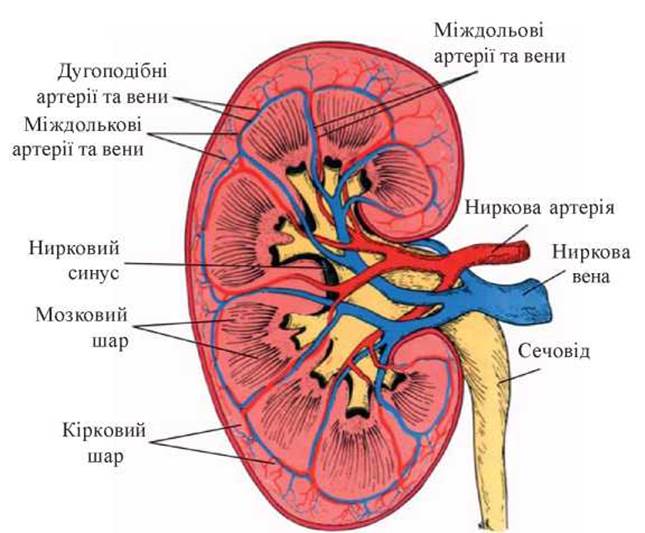
Рис.10. Схема кровопостачання нирки (за Г. Маждраковым и соавт., 1980).
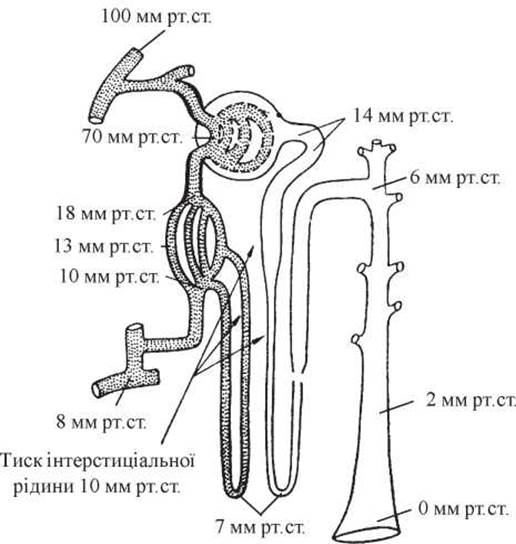
Рис. 11. Артеріальний тиск у різних відділах нефрона (за А.С. Guyton, 1971).
Досить складною за будовою є і Нервова система нирок. У нирки проникає більша (порівняно з іншими внутрішніми органами) кількість нервових волокон. Вони відходять від черевного сплетення на рівні XII грудного - II поперекового хребців і утворюють у воротах нирки ниркове сплетення (plexus radialis), у якому виділяють верхні і нижні аортониркові вузли та дрібні ниркові вузли. Ниркове сплетення тісно пов’язане з аортальним, сонячним сплетеннями і поперековими вузлами симпатичного стовбура. Тораколюмбальний Симпатичний стовбур забезпечує нирку великою кількістю вазоконстрикторних волокон, у той час як кількість парасимпатичних волокон значно менша (Блукаючий нерв надсилає у нирку лише невелику кількість своїх гілочок). Таким чином, відповідний ситуації вазомоторний статус нирки переважно підтримується за рахунок змін вазоконстрикторних впливів адренергічних нервів.
Адренергічна іннервація домінує у корі, незначну кількість норадреналіну знайдено у зовнішньому медулярному шарі, однак його менше, ніж у внутрішньому. Адренергічні нерви супроводжують міжчасточкові, дугоподібні артерії й аферентні артеріоли, що зумовлює можливість їх вазоконстрикції за рахунок скорочення гладеньком’язових клітин судинної стінки. Цих нервів не знайдено у гломерулах, еферентних артеріолах, канальцях.
Холінергічні волокна переважно розміщуються у ділянці еферентних артеріол юкстамедулярних нефронів і сфінктерів vasa recta. Таким чином, медулярний Кровообіг, а також ефективний контроль за внутрішньоклубочковим тиском у юкстагломерулярних нефронах пов’язані виключно з цими вазодилататорними волокнами.
Ниркова гемодинаміка переважно контролюється альфа-адренорецепторами, а виділення реніну - бета-адренорецепторами. Бета-адреностимуляція викликає розширення ниркових судин, альфа-адреноблокада - їхнє звуження. Стимуляція альфа- і бета-адренорецепторів супроводжується викидом вазопресину, еритропоетину. Інтенсивна стимуляція симпатичних нервів нирки викликає зменшення кровообігу і фільтрації сечі, помірна - зменшення кровообігу і збільшення фільтрації сечі. Принагідно вкажемо, що повністю денервована нирка здатна підтримувати на належному рівні такі показники, як КФ, ренальний плазмотік, що узгоджується із концепцією про високий ступінь автономності ренальної гемоциркуляції.
Нирки беруть активну участь у регуляції сталості внутрішнього середовища за рахунок впливу на об’єм, іонну концентрацію й осмотичний тиск позаклітинної рідини, кислотно-лужну рівновагу (КЛР), Метаболізм білків, жирів, вуглеводів, вітаміну Д3, еритропоез, системний та регіонарний кровобіг.
Сучасне розуміння механізмів утворення сечі включає в себе процеси ультрафільтрації в клубочках, реабсорбції та секреції в канальцях, а також синтез ниркою певних сполук.
Початковий процес сечоутворення - ультрафільтрація плазми крові. На даний час гломерулярний фільтр розглядається як селективна система з бар’єрами зростаючої щільності. До його складу входять ендотелій капілярів, щільний шар БМ капілярів клубочків, щілиноподібна діафрагма подоцитів. У механізмі ультрафільтрації особливого значення надають фізико-хімічному стану БМ (величині її негативного заряду) та стану ниркового капілярного кровобігу, який у нормі формує ще один (функціональний) бар’єр із високомолекулярних білків плазми - так званий шар концентрації-поляризації. Зворотні ізометричні зміни в’язкості колоїдів зовнішнього шару БМ, глікокаліксу ендотелію капілярів і подоцитів, які визначають розмір пор фільтрації, стерична відповідність форми речовини, яка фільтрується, і пор для її фільтрації, зміни величини заряду БМ (уроджені або набуті), порушення внутрішньоклубочкового тиску зумовлюють швидкість гломерулярної фільтрації і склад фільтрату в нормі та при патології. Ультрафільтрат практично не містить білків, має аналогічну з плазмою концентрацію електролітів, глюкози, сечовини, креатиніну. У ньому можна знайти лише невелику кількість білків із масою 12-21 кілодальтон (бета-2-мікро-глобулін, лізоцим, ретинозв’язувальний білок). Загальна концентрація альбумінів в ультрафільтраті складає 1 мг/л, у плазмі крові - 40-50 г/л; неушкоджена гломерулярна мембрана пропускає в ультрафільтрат не більше 0,1 % альбумінів крові. Об’єм КФ складає в середньому 180 л/24 год, а загальний вміст білка у фільтраті - 36 г/24 год. Принагідно зазначимо, що загальна добова фільтрація рідини через мембрани всіх капілярів тіла, крім мембран клубочків нирок, складає лише 4 л, і профільтрована рідина повертається в центральний кровотік по лімфатичних судинах. Вкажемо також, що швидкість гломерулярної фільтрації є величиною відносно стабільною і майже не залежить від коливань системного AT у фізіологічно допустимих межах. Це зумовлюється наявністю добре виражених авторегуляторних механізмів, серед яких провідне місце, як уже згадувалося, займає юкстагломерулярний апарат. У той же час клубочок (його судинний і мезангіальний компоненти) піддається впливу інших продукованих локально і системно фізіологічно активних речовин - вазопресину, брадикініну, ПГ, норадреналіну, ацетилхоліну тощо.
Вирішальним фактором, який забезпечує процес ультрафільтрації в клубочках, є різниця між величиною гідростатичного тиску, який створюється роботою серця, і протидійними силами. До останніх належать онкотичний тиск плазми крові (1626 мм рт.ст.) і тиск рідини безпосередньо в капсулі клубочка (8-15 мм рт.ст.).
Таким чином, різниця, яка створює ефективний фільтраційний тиск, знаходиться в межах 8-12 мм рт.ст., а сумарний об’єм КФ залежить від кількості функціонуючих клубочків, стану клубочкового кровобігу, величини коефіцієнта проникності капілярної стінки.
Просвіт клубочкової капсули безпосередньо переходить у проксимальний відділ канальця (його довжина складає 13-14 мм при загальній довжині канальця 50 мм). Цей відрізок канальця вкритий одношаровим кубічним епітелієм, клітини якого містять базально розміщене Ядро, велику кількість мітохондрій і гранул рибонуклеїнової кислоти. Поверхня клітин, яка обернена до просвіту канальця, покрита щетинками, що значно збільшують площу всмоктувальної поверхні. У зоні щетинок зосереджена велика кількість лужної фосфатази, АТФ-ази, 5-нуклеотидази, а в численних мітохондріях - дегідрогеназ, діафораз, гідролаз та інших окиснювальних ферментів.
Ультраструктура і ферментохімія клітин головного відділу канальців пристосовані до виконання колосальної за об’ємом функції реабсорбції білка, води, глюкози, амінокислот, електролітів, вітамінів. Вважають доведеним, що весь профільтрований у клубочках білок реабсорбується в проксимальному відділі канальця (енергозалежна вакуольно-лізосомальна система реабсорбції). Звідси - важлива роль ряду інфекцій, інтоксикацій або генетично детермінованих порушень у виникненні гострої або хронічної блокади цієї системи, наслідком чого є ПУ, розвиток гострої ниркової недостатності (ГНН).
Структура епітелію тонкого сегмента петлі Генле пристосована до максимального зменшення фільтраційного заряду води та її «пасивної» резорбції. Найбільш вузька частина канальця в ділянці петлі Генле покрита сплощеним епітелієм зіркоподібної форми. Ці клітини майже не містять окиснювальних ферментів, кількість мітохондрій у них обмежена, проміжки між клітинами заповнені скріплювальною тканиною. Така структура епітелію зумовлює обмежену реабсорбцію в цьому відділі нефрона і пов’язана з функціонуванням так званого протиточно-множувального механізму концентрування сечі (рис. 12).
Принцип роботи цього механізму сформульований ще у 50-і роки (H. Wirz et al., 1951) і від того часу уявлення про нього змінилися мало. Здатність нирки концентрувати сечу тісно пов’язана зі структурними особливостями нефрона, розміщенням петлі Генле, збиральних проток та кровоносних судин.
До складу петлі Генле входять два паралельно розміщені коліна з протилежним напрямком руху сечі в них. Кровоносні судини мозкового шару - vasa recta - повторюють згини петлі, а потік крові у низхідному та висхідному відрізках судин теж протилежно направлений. Уся ця структура нефрона технічно нагадує протиточно-множувальний механізм, який сприяє створенню більш високих концентрацій речовини за рахунок невеликої концентраційної різниці в кожному із колін при проходженні рідини вздовж усієї системи. В юкстамедулярних нефронах, які розміщені на межі кіркового і мозкового шарів, обидва коліна петлі Генле та судинні гілочки, які їх оточують, проходять через мозковий шар до його пограничних відділів. Тут вони здійснюють поворот і прямують до кіркового шару. Завдяки інтенсивній реабсорбції натрію в ділянці петлі Генле та сповільненню кровобігу у vasa recta в інтерстиціальній рідині цього шару створюється висока осмотична концентрація натрію. Ця величина зростає в напрямку від кіркового шару до вершини сосочка.
Таким чином, механізм концентрування сечі наступний: при проходженні проксимального відділу канальця ізотонічний фільтрат у зменшеному об’ємі надходить у петлю Генле. На цій ділянці інтенсивна реабсорбція натрію не супроводжується реабсорбцією води, оскільки стінка канальця малопроникна для неї. Результатом цього явища є розведення сечі в просвіті нефрона і підвищена концентрація натрію в інтерстиції. Розведена сеча в дистальному канальці втрачає надлишок рідини і стає рівноізотонічною плазмі. Зменшений об’єм ізотонічної сечі надходить у збиральну систему, яка розташована у мозковому шарі і навколо якої існує високий осмотичний тиск внаслідок підвищеної концентрації натрію в інтерстиції. У збиральних трубках під впливом антидіуретичного гормону продовжується зворотне всмоктування води відповідно до концентраційного градієнта. Vasa recta, які проходять у мозковому шарі, функціонують як протиточно-обмінні структури - вони забирають на шляху до сосочків натрій і віддають його до свого повернення в кірковий шар. Таким чином, у глибині мозкового шару підтримується високий вміст натрію, який забезпечує резорбцію води зі збиральної системи, тобто процес концентрування сечі. Енергія, необхідна для виконання нирками такої осмотичної роботи, є досить великою. Так, для виведення нирками за 24 години 1 л сечі необхідно затратити 848 калорій.
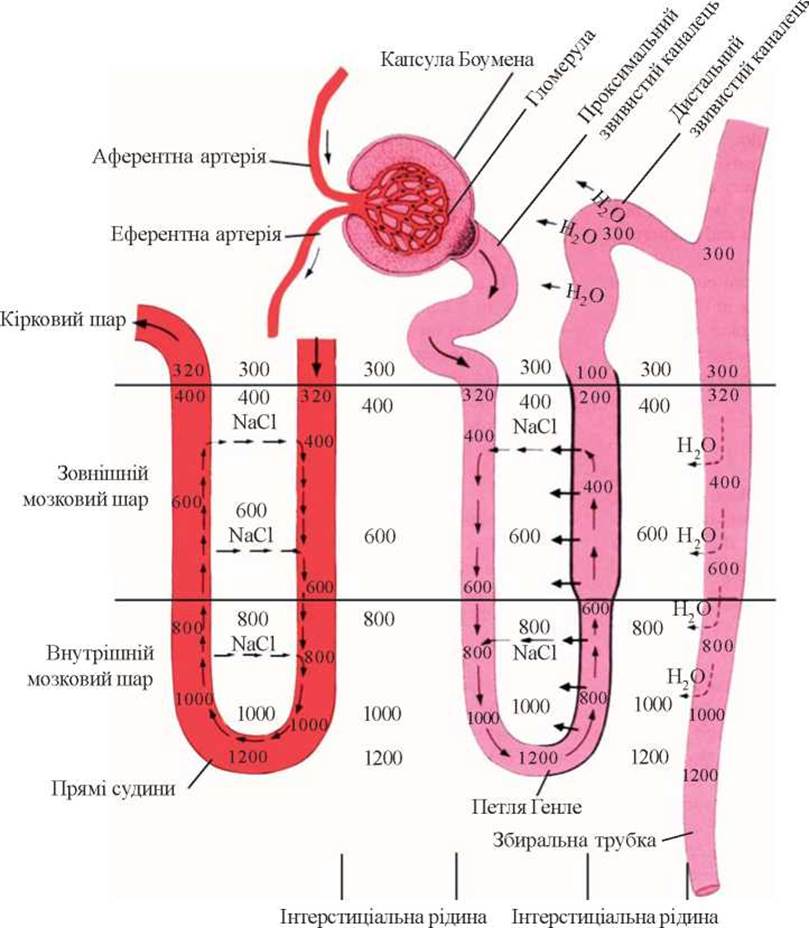
Рис. 12. Схема протиточно-множувального механізму концентрування сечі.
Дистальний відділ канальця вкритий кубічним епітелієм, клітини якого містять апікально розміщене ядро, велику кількість сигароподібних мітохондрій, гранули рибонуклеїнової кислоти та дрібні вакуолі. Останні нагадують міхурці соляної кислоти в залозистому епітелії слизової шлунка. Епітелій дистального відділу канальця багатий на амінокислоти, основні та кислі білки рибонуклеїнової кислоти, Полісахариди та реактивні SH-групи; для нього характерний високий вміст гідролітичних, гліколітичних ферментів і ферментів циклу Кребса. Всі ці структури забезпечують належну біохімічну активність, яка пов’язана з необхідністю підтримувати на високому рівні іонний обмін, синтез амонію та ін. Зі структурно-функціональними особливостями дистальних канальців, їх підвищеною чутливістю до гіпоксії пов’язують і їх першочергове ураження (аж до некрозу) при дії токсичних продуктів кислої сечі.
Загалом, сумарна довжина канальців сягає 100 км, а поверхня канальцевого епітелію - 50-60 м2.
У ділянці збиральних трубок кубічний епітелій поступово набуває циліндричної форми (світлі і темні клітини з добре розвинутим базальним лабіринтом та високою проникністю для води); кількість клітинних органел у ньому незначна. З темними клітинами пов’язують процес секреції іонів водню. Нефроцити цього відділу нефрона секретують ПГ, а клітинні мембрани мають властивість змінювати свою проникність під дією антидіуретичного гормону.
Таким чином, специфіка будови епітелію канальців, висока ферментативна активність внутрішньоклітинних структур забезпечують вибіркову реабсорбцію або секрецію тих чи інших елементів для Підтримання гомеостазу (рис. 13).
Зазначимо, що виділення окремих кінцевих продуктів обміну та введених ззовні органічних речовин відбувається переважно за рахунок канальцевої секреції. Так, шляхом секреції клітинами проксимального відділу нефрона у просвіт канальця виділяються жовчні кислоти, сечова кислота, цАМФ, адреналін, ацетилхолін, гістамін, серотонін, діодраст, індометацин, пеніцилін, фуросемід, етакринова кислота.
У забезпеченні водно-електролітного гомеостазу, зокрема гомеостазу натрію і калію, ниркам належить провідна роль. Натрій і калій - основні електроліти, розподіл яких у внутрішньо- і позаклітинних рідинах є вирішальним фактором для підтримання нормальної осмотичної концентрації позаклітинної рідини. Концентрація натрію в плазмі і позаклітинній рідині складає 130-150 ммоль/л, у внутрішньоклітинній - 1113 ммоль/л, калію - відповідно 4,0-4,5 ммоль/л і 140-150 ммоль/л. У нормі загальний осмотичний тиск внутрішньо- і позаклітинної рідин однаковий, рух рідини через клітинні мембрани відсутній, і перерозподіл рідинних просторів організму виникає лише при зміні концентрації натрію в позаклітинній рідині. Його нормальна концентрація в позаклітинній рідині регулюється за рахунок механізму спраги, який забезпечує надходження рідини в Організм, і секреції антидіуретичного гормону, який регулює виділення цієї рідини з організму.
Роль нирок у підтриманні балансу натрію полягає в контролі за процесом виділення натрію з сечею. Ці механізми контролю надзвичайно складні і багатогранні. Вільно фільтруючись у клубочках, натрій послідовно реабсорбується в усіх сегментах ниркових канальців залежно від потреб організму. Таким чином, регуляцію балансу натрію розглядають із позицій співвідношень величин його КФ та канальцевої реабсорбції (клубочково-канальцевий баланс, Джеймс А. Шейман, 1999). Дані літератури (M. Shapiro et al., 1986; H.J. Reineck et al., 1993) свідчать, що при всій важливості швидкості КФ основним фактором у забезпеченні натрієвого гомеостазу є все-таки канальцева реабсорбція. Цей процес контролюється альдостероном, натрійуретичними пептидами (передсердним гормоном та його аналогами), фізичними факторами, які діють у ділянці проксимального канальця (різниця між гідростатичним і онкотичним тиском у капілярах канальця і його просвіті, величина та швидкість перитубулярного кровобігу, проникність капілярної стінки), перерозподілом внутрішньониркового кровобігу, кортикостероїдами, естрогенами, гормоном росту, інсуліном, прогестероном, катехоламінами, калікреїн-кініновою системою, ПГ (M. Cogan, 1990; T. Ferris, 1993). Із позаниркових факторів найбільш важливу роль у процесі реабсорбції натрію надають альдостерону у складі ренін-ангіотензин-альдостеронової системи (РААС), хоча в кількісному вираженні він регулює лише 2 % загальної кількості реабсорбованого натрію. Річ у тім, що точкою його дії є головні клітини збиральних трубок кори нирок - ділянка нефрона, до досягнення якого вже реабсорбувалося близько 90 % натрію; тобто на цій ділянці остаточно вирішується питання реабсорбції кількості натрію для підтримання гомеостазу системи.
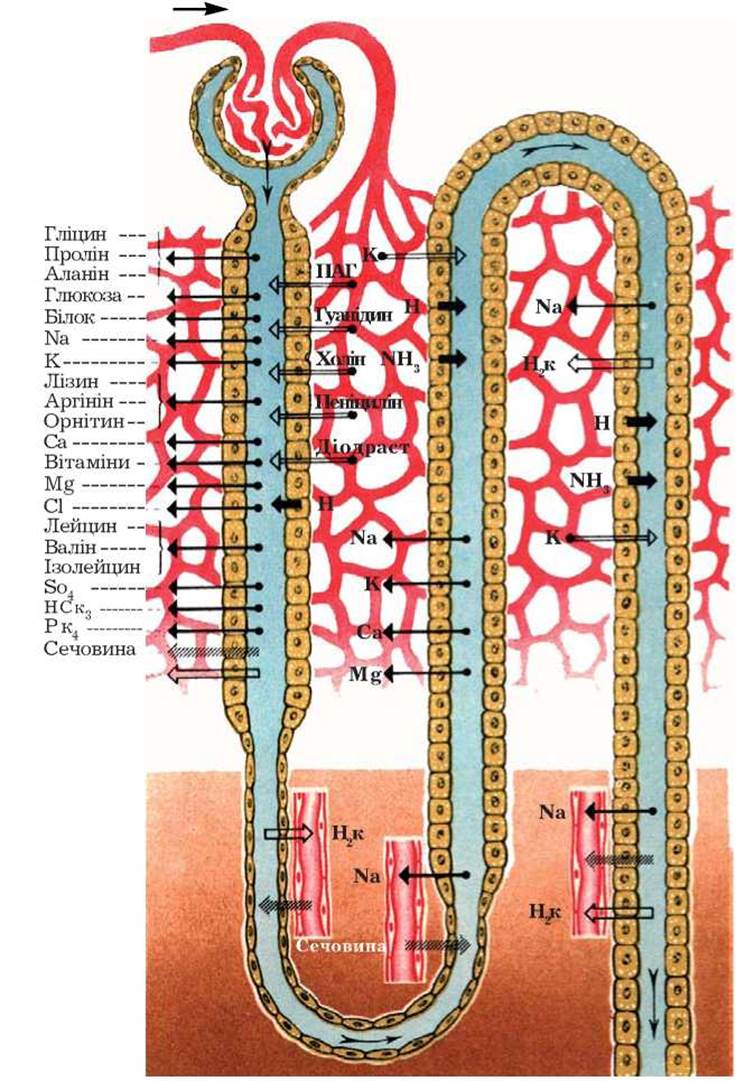
Рис.13. Схема локалізації транспорту речовин у нефроні. Стрілка, направлена в просвіт канальця, - секреція, із просвіту канальця - реабсорбція (Ю.В. Наточин, 1974).
Нормальна нирка фільтрує за добу близько 23 моль Cl-. Зі сечею екскретується тільки 1 % від цієї кількості, а 99 % профільтрованих іонів хлору реабсорбується у канальцях - 50-60 % у проксимальних, 5-10 % - у дистальних відділах, 2030 % - у товстому сегменті висхідного коліна петлі Генле, 2-5 % - у збиральних трубках. Механізми реабсорбції хлору гетерогенні - доведено існування спеціалізованих «хлорсекретуючих клітин», які, власне, і є модуляторами процесу його реабсорбції.
Позаклітинний вміст калію теж контролюється досить строго, переважно альдостероном. Розподіл калію підтримується, головним чином, Na+, K+АТФ-азою - універсальним транспортером у складі всіх клітинних мембран. Зростання концентрації калію в плазмі крові підвищує секрецію альдостерону, зменшення - знижує її, і цей зворотний зв’язок є надзвичайно важливим. До кінця товстого відділу висхідного коліна петлі Генле реабсорбується в середньому 90 % калію, а остаточне рішення щодо затримки або виділення нирками калію реалізується на рівні збиральних трубок: при високому вмісті калію в їжі ці структури секретують його, при низькому - секреція відсутня. Крім цього, в регуляції гомеостазу калію беруть участь і позаниркові фактори - рівень інсуліну в крові, стан бета-адренергічної системи. Механізм, за допомогою якого інсулін регулює вхід калію в клітини, точно не визначений. Існує думка, що інсулін може активувати мембранні Na+, Н+-насоси. З іншого боку, гіперкаліємія стимулює, а гіпокаліємія інгібує виділення інсуліну. Катехоламіни посилюють процес входження калію в клітину шляхом активації бета-адренергічних рецепторів; стимуляція альфа-адренорецепторів супроводжується протилежним ефектом. Не можна не згадати і про ацидоз - стан, при якому відбувається вихід калію з клітини. Зсув рН крові на 0,1 супроводжується змінами сироваткової концентрації калію на 0,6 ммоль/л.
Ще більш складною є система регуляції кальцієво-фосфорного обміну, що пов’язано з надзвичайно важливою роллю кальцію в гомеостазі (В.В. Коломиец, 1990; В.О. Бобров і співавт., 1994). Цей іон бере активну участь у майже всіх внутрішньоклітинних процесах, генерації електричного потенціалу клітини, активації багатьох внутрішньоклітинних ферментів, утворенні нових біологічних структур, процесах Запліднення, секреції, нервово-м’язової провідності та м’язового скорочення, мембранної проникності, проміжному обміні тощо. Від концентрації кальцію залежать сила скорочень кардіоміоцитів, тонус судинної стінки та бронхів, рухова активність матки, шлунково-кишкового тракту, сечоводів, секреція інсуліну, гормонів гіпофіза та інших ендокринних залоз, агрегація тромбоцитів (В .А. Бобров и соавт., 1992; R.D. Bukowsky et al., 1991; J.T. Repke et al., 1991; K.B. Knight et al., 1992). Тому концентрація кальцію в крові, точніше, іонізованого кальцію (В.Н. Титов и соавт., 1992), який і визначає його фізіологічну роль, є одним із найбільш стабільних параметрів електролітного гомеостазу.
Із трьох різних фракцій кальцію, які є у сироватці крові (іонізована, 44 % від загальної концентрації кальцію, комплексна (хелатна) - 10 % та білковозв’язана - 46 %) тільки іонізований кальцій та його хелатні сполуки здатні проникати крізь клубочкові капіляри у первинну сечу. Вкажемо, що іонізований кальцій сироватки крові є не тільки важливим об’єктом регуляції в системі гомеостазу кальцію, а й сам служить фактором, який регулює секрецію кальційрегулювальних гормонів.
Найбільш важливе значення в регуляції концентрації кальцію в крові відіграють паратгормон, Кальцитонін і 1,25 (ОН)2Д3.
Паратгормон (гормон паращитоподібних залоз) підвищує реабсорбцію кальцію в дистальних канальцях, знижує канальцеву реабсорбцію натрію, фосфатів і ряду амінокислот, інгібує секрецію водневих іонів та натріє-водневий обмін, що супроводжується бікарбонатурією. Поряд із локально синтезованим туморонекротичним фактором-альфа та інтерлейкіном-1, він сприяє також резорбції кальцію з кістки шляхом активації діяльності остеокластів, не впливаючи суттєвим чином на процеси абсорбції кальцію в кишечнику. При прогресуванні ниркового процесу у кістковому мозку розвивається помітний фіброз, і поступово виявляється гістологічна картина фіброзного оститу. Однак паратгормон водночас активує діяльність і остеокластів (Ш. Массри, 1999). Таким чином, прояви надлишку паратгормону в кістках у пацієнтів із ХНН включають у себе підвищену кількість остеокластів і остеобластів, посилену остеокластичну кісткову резорбцію, збільшення гаверсових порожнин, ендостальний фіброз, накопичення волокнистої кісткової Тканини і волокнистих остеоїдів.
На противагу паратгормону, кальцитонін (гормон міжфолікулярних клітин щитоподібної залози) знижує концентрацію кальцію в плазмі, попереджуючи таким чином виникнення гіперкальціємії. Будучи функціональним антагоністом паратгормону, кальцитонін шляхом впливу на специфічні рецептори зовнішньої мембрани зменшує кількість і активність остеокластів у кістковій тканині, інгібує остеоцитний остеоліз, індукований паратгормоном. Кальцитонін має також фосфатуричний ефект за рахунок пригнічення процесу реабсорбції фосфатів у проксимальних відділах канальців.
За вмістом вказаних гормонів у сироватці крові можна виділити дві групи пацієнтів: перша - з нормальним рівнем лужної фосфатази, високими значеннями кальцитоніну і порівняно низьким вмістом паратгормону, друга - з високими рівнями лужної фосфатази, паратгормону і низьким - кальцитоніну. Ці дані свідчать про існування механізму негативного кореляційного зв’язку між паратгормоном і кальцитоніном, який реалізується, ймовірно, опосередковано через концентрацію іонізованого кальцію у крові (С.И. Рябов и соавт., 2000).
Нарешті, важлива роль у забезпеченні гомеостазу кальцію належить 1,25 (ОН)2Д3 - найбільш метаболічно активній формі вітаміну Д3. У нормі вітамін Д3 утворюється в шкірі або надходить в організм з їжею. Частина цієї біологічно неактивної речовини у мікросомальній та мітохондріальній фракціях печінкових клітин перетворюється у 25-гідроксикальциферол (25(ОН)Д2), із якого у проксимальних звивистих канальцях нирок утворюється 1,25(ОН)2Д3. Продукцію 1,25(ОН)2Д3 одночасно стимулюють паратгормон (цАМФ-залежний шлях) і кальцитонін (шляхом підвищення активності 1-альфа-гідроксилази).
Метаболіти вітаміну Д3, які у здорової людини утворюються лише в нирках, зумовлюють переважно екстраренальні ефекти - підвищення вмісту кальцію в крові за рахунок посилення процесу резорбції кістки, процесу абсорбції кальцію в кишечнику і зменшення екскреції кальцію зі сечею. Вітамін Д3 змінює мінералізацію кісткової тканини різними шляхами - впливом на синтез і дозрівання колагену, прямою стимуляцією і/або підвищенням рівнів кальцію і фосфору в міжклітинній рідині, яка оточує кістку (Ш. Массри, 1999).
Зазначимо, що поряд із даними про біологічну роль вітаміну Д3 у регуляції мінерального обміну, останнім часом з’явилися повідомлення про вплив його активних метаболітів на стан імунокомпетентних клітин (Ф.И. Руснак и соавт., 1997). Доведено, наприклад, що при інфекційно-алергічних захворюваннях практично завжди виявляють експресію лімфоцитарних рецепторів до кальцитріолу, тоді як серед здорових людей така експресія визначається лише у 17 % випадків.
Однак роль нирок у підтриманні кальцієвого гомеостазу цим не вичерпується.
Важливе значення в цьому процесі відіграє участь нирки в метаболізмі кальційрегулювальних гормонів (Дж. Леман и соавт., 1987; J.H. Laragh et al., 1992). За нормальних умов паратгормон і кальцитонін безперервно фільтруються в ниркових клубочках, і ультрафільтрат потрапляє у просвіт проксимальних канальців. Там ці гормони гідролізуються до амінокислот, і останні всмоктуються в кров для їх нового синтезу. Активна форма вітаміну Д3 , як і інші стероїдні гормони, теж поступово інактивується в нирках, що створює передумови для підтримання її постійного рівня в крові за рахунок зміни швидкості процесу інактивації. Стан різко змінюється при виникненні ниркової недостатності (НН) (H. Saha et al., 1991; Y. Tsucamoto et al., 1991). Зменшення маси нефронів призводить до зниження швидкості фільтрації гормонів, їх інактивації, наступного накопичення в крові з одночасним зменшенням процесу синтезу 1,25(ОН)2Д3. У результаті розвиваються важкі порушення обміну кальцію, виникають Остеомаляція, фіброзний остеїт, остеосклероз.
Зазначимо і несприятливий вплив на баланс кальцію і розвиток скелетних аномалій ацидозу. Гострий ацидоз викликає значну втрату скелетом розчинного у кислоті карбонату кальцію. За стабільної НН з притаманною їй стійкою затримкою іонів водню Скелет забезпечує запаси буфера за рахунок зменшення запасів карбонату кальцію.
Уже на ранній стадії порушення функції нирок у результаті зниження фільтраційного заряду фосфору розвивається гіперфосфатемія, яка супроводжується реципрокним зменшенням у крові кількості іонізованого кальцію. Останнє індукує секрецію паратгормону, фосфатурію. Таким чином, ціною гіперсекреції паратгормону і зростання резорбції кістки рівень фосфору в крові повертається до норми. При подальшому зниженні КФ і підвищенні рівня фосфору в крові повторно індукується секреція паратгормону.
Загальна кількість магнію в організмі становить близько 2000 мекв або 25 г. 60 % цієї кількості знаходиться у кістках, 20 % - у м’язах, 20 % - у інших органах і тканинах, позаклітинній рідині.
Нирки фільтрують близько 2 г магнію/добу, а його екскреція складає лише 100-130 мг/добу. Встановлено, що між реабсорбцією кальцію і магнію існує реципрокний зв’язок - Введення одного з цих електролітів знижує реабсорбцію іншого. Застосування мінералокортикоїдів, алкоголю, глюкози збільшує екскрецію магнію, причому провідна роль у цьому процесі належить висхідній частині петлі Генле; роль термінальних сегментів нефрона мінімальна.
Основним механізмом регуляції ниркою КЛР є канальцева секреція іонів Н+, яка забезпечує реабсорбцію бікарбонату та еквівалентної кількості Na+. Максимальна реабсорбція бікарбонату (80-90 %) відбувається у проксимальному канальці; 5 % бікарбонату реабсорбується у петлі Генле, 3-4 % - у дистальних звивистих канальцях. Секреція іонів Н+ відіграє важливе значення і для виділення ниркою кислих продуктів (фосфатів, аміаку). Поєднанням цих двох процесів - реабсорбції бікарбонатів і секреції Н+ - забезпечується постійність pH крові (у нормі - 7,35-7,43). Певна роль у регуляції pH крові належить альдостерону (сприяння секреції Н+ шляхами підвищення негативного заряду БМ і безпосередньої стимуляції Н+ АТФ-ази), а також паратгормону, про що згадувалося вище. Відзначимо, що реакція нирок на зміни КЛР крові відбувається набагато повільніше, ніж відповідна реакція зі сторони органів Дихання.
Наводимо таблицю 1, у якій вказані величини фільтрації, реабсорбції й екскреції основних неорганічних та органічних речовин.
Значно розширилися і наші знання про активну роль нирок у підтриманні фізіологічного стану системи гемостазу.
Нормальні агрегатні властивості крові - вкрай необхідна умова для виконання нирками своїх функцій. У нормі нирки продукують та виділяють у кров та сечу ряд факторів-прокоагулянтів (VII, VIII, IX, X, тромбопластин), а також синтезують та екскретують такі протизгортальні фактори, як гепарин, урокіназа, тканинний активатор плазміногену. Нирки поглинають і катаболізують частину фібриногену і XIII фактор, який бере участь у синтезі колагену БМ капілярів клубочків. За умов патології при депозитуванні імунних комплексів і/або їх утворенні in situ в клубочках, при розвитку запалення з посиленою транссудацією білків-прокоагулянтів, з однієї сторони, втрачається функціональний вклад нирок у гемостаз, з іншої - виникає процес локальної активації згортальної системи крові з внутрішньосудинною гіперкоагуляцією. Така локальна гіперкоагуляція може за певних умов перейти у дисемінований процес.
Важливою є і роль нирок у регуляції гемопоезу. Гіпохромна нормоцитарна анемія - класичний симптом порушення функції нирок. її Патогенез складний і включає в себе різноманітні ланки - посилений гемоліз, стан гіперкатаболізму, мікрокровотечі (Г.Д. Шостка, 1997), серед яких провідну роль відводять порушенню продукції нирками еритропоетину.
Еритропоетин - гліколізований поліпетид із молекулярною масою 34 000, до складу якого входять 165 амінокислот. Його карбогідратна частина відповідає за збереженість біологічної активності гормону в організмі, а безсіалова швидко руйнується гепатоцитами. Основним стимулом для його утворення є зниження рО2 безпосередньо в капілярах ниркової паренхіми, точніше, у венозному кінці капіляра. Власне тому гіперпродукцію еритропоетину пов’язують не тільки з патологією нирок, але і з патологією крові, серцево-судинної і дихальної систем.
Першим етапом каскадної реакції, яка призводить до посиленого синтезу еритропоетину, є підвищене надходження кальцію у простагландинпродукуючі клітини ниркових структур - виявлено прямий стимулювальний вплив ПГ Е2 на еритропоез у кістковому мозку. Продукцію еритропоетину стимулюють також гормони кори надниркових залоз, тиреоїдин, андрогени.
Рецептори-стимулятори продукції еритропоетину (сенсорні клітини) завжди повинні мати тісний і постійний контакт із кров’ю. Такою зоною в нирках є кіркова речовина і зовнішня частина мозкового шару, в яких за допомогою механізму авторегуляції забезпечується постійність кровобігу в досить широкому діапазоні АТ (від 70 мм рт.ст. до 200 мм рт.ст.). За останніми даними, ці рецептори-стимулятори розміщені в ендотеліальних клітинах перитубулярних капілярів проксимального відділу канальців кіркового і мозкового шарів. Там же, найбільш імовірно, і продукується еритропоетин. За умови вираженої гіпоксії еритропоетин може продукуватися та інактивуватися гепатоцитами і клітинами Купфера.
У кістковому мозку еритропоетин сприяє переходу уніпотентних еритроїдних комітованих попередників еритропоезу в еритрон, початковою клітиною якого є проеритробласт, кінцевою - еритроцит.
Виділяють декілька напрямків дії еритропоетину на клітинному рівні: стимуляція процесів проліферації і дозрівання ранніх проміжних форм еритропоетинчутливих клітин, вкорочення інтермітотичного періоду, прискорення дозрівання нормобластів і кістковомозкових ретикулоцитів, виключення одного або декількох мітотичних поділів, зменшення частки так званого «неефективного» еритропоезу.
Іншою важливою причиною анемії при патології нирок є гемоліз внаслідок підвищеного руйнування еритроцитів у морфологічно змінених (відкладання фібрину, дистрофічні зміни ендотелію) артеріолах і капілярах. Проходячи через такі змінені мікросудини, еритроцити травмуються, втрачають свою форму і швидко гемолізуються.
Вкажемо також на змінений при НН метаболізм заліза - його зменшене вживання при перебуванні на малобілковій дієті, токсичний вплив на синтез гему, метаболізм порфірину алюмінію діалізної води та паратгормону (Г.Д. Шостка, 1999; A. Lach et al., 1995).
В останні роки з’явилися дані про негативний вплив на еритропоез підвищеного сироваткового вмісту середньомолекулярних пептидів (СМП) (Р.З. Исмагилов и соавт., 1998; D.S. Rao et al., 1993).
Порівняно рідко при захворюваннях нирок виникає еритроцитоз. Такий стан гемопоезу характерний для пухлин нирок, зокрема гіпернефроїдного раку, ниркових кіст, гідронефрозу, полікістозу. Пояснення цьому знаходять у підвищеній продукції еритропоетину при вказаних формах патології.
Транспорт сечі на заключному етапі виділення (з нирки в сечовий міхур) - теж активний процес (Г. Маждраков и соавт., 1980). Рух сечі забезпечується скороченням м’язових волокон, які розміщені в стінці сечових шляхів, і цей активний процес починається на рівні ниркових мисок.
У мускулатурі чашечок і мисок розрізняють чотири дрібних м’язи: m. levator fornicis, m. sphinkter fornicis, m. longitudinalis calycis, m. sphincter calycis. Діяльність цих м’язів строго скоординована, що і забезпечує нормальний пасаж сечі. Під час так званої діастолічної фази m. levator fornicis та m. sphinkter fornicis розслабляються, у той час як верхня частина m. longitudinalis calycis знаходиться у скороченому вигляді, а m. sphincter calycis відмежовує порожнину чашечок від мисок. У результаті цього в просторі навколо сосочків створюється від’ємний тиск, який сприяє «висмоктуванню» сечі з нирки. Услід за цим настає систола чашечок, протягом якої відбувається скорочення m. levator fornicis та m. sphinkter fornicis при одночасному розслабленні m. longitudinalis calycis та m. sphincter calycis. Наповнення чашечок (діастола) триває 30-60 с, спорожнення (систола) - 1-3 с.
За допомогою такого механізму сеча з нирки невеликими порціями впорскується в миску. Поки відбувається наповнення розслабленої миски, найбільш дистальна частина її шийки залишається закритою. Надалі при одночасному розкритті шийки миски і скороченні м’язів її стінки певна порція сечі надходить у проксимальний відділ сечовода.
У сечоводі утворюється наповнене сечею гострокінцеве «веретено». Це «веретено» рухається по сечоводу зі швидкістю 2,5-3 см/с. Одночасно по сечоводу проходить лише одне «веретено», яке на заключному етапі виштовхується в сечовий міхур. За необхідності (підвищений діурез) збільшуються довжина і радіус «веретен» та зменшуються інтервали між їх виникненням; при дуже високому діурезі (наприклад, нецукровий діабет) виникає фізіологічна гіпотонія сечоводів.
У міру Наповнення сечового міхура сеча шляхом вільного сечовипускання виділяється через уретру назовні.
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.