Гістологія людини - О.Д. Луцик 2003
Цитологія
Клітинна оболонка. Цитоплазма
Клітинна оболонка (плазмолема, plasmolemma) В основі будови плазмолеми знаходиться елементарна біологічна мембрана. Структуру останньої описує рідинно-мозаїчна модель Сінгер-Нікольсона. Суть її така: молекули фосфоліпідів, контактуючи своїми гідрофобними кінцями і відштовхуючись гідрофільними, утворюють суцільний подвійний ліпідний шар, у який частково або повністю втоплені молекули білків (переважно глікопротеїнів). Молекули білків, які пронизують усю товщу біліпідного шару або значною мірою втоплені в нього, - це так звані інтегральні білки; ті ж білки, які розміщені лише на поверхні ліпідів, називаються периферійними або адсорбованими (рис.1.3). Положення білкових молекул не є жорстко лімітованим - залежно від функціонального стану клітини може відбуватися їхнє взаємне переміщення у площині біліпідного шару. Ця Мінливість і подібна до мозаїки топографія макромолекулярних комплексів поверхні клітини дала назву рідинно-мозаїчній моделі біологічної мембрани. Лабільність (плинність) структур плазмолеми залежить від вмісту у її складі молекул холестерину: чим вищий вміст холестерину, тим легше переміщуються макромолекулярні білкові комплекси у біліпідному шарі. Серед білків плазмолеми існує певна спеціалізація: є структурні, ферментні, транспортні, рецепторні молекули. Однією з важливих умов нормального функціонування біологічної мембрани є збереження принципу замкнутості (відсутності розривів) біліпідного шару.
Вуглеводні компоненти молекул глікопротеїнів та гліколіпідів плазмолеми, випинаючись над зовнішньою поверхнею клітинної мембрани, формують, так звану, надмембранну зону, або глікокалікс. Олігосахаридні ланцюги глікокаліксу є своєрідною "візитною карткою" клітини. За їхньою участю здійснюється взаєморозпізнавання клітин та взаємодія з мікрооточенням. Кожному різновиду клітин притаманна особлива послідовність моносахаридних залишків у складі поверхневих олігосахаридних ланцюгів глікополімерів, свій унікальний набір і цитотопографія вуглеводних детермінант.
З боку внутрішнього вмісту клітини з мембраною контактує, так звана, внутрішня (підмембранна) пластинка, або кортикальний шар цитоплазми. Це найбільш в'язка частина цитоплазми, багата на мікрофіламенти і мікротрубочки, що утворюють високоорганізовану сітку і за участю яких здійснюються, зокрема, переміщення інтегральних білків плазмолеми, забезпечуються цитоскелетні та локомоторні функції клітини, реалізуються процеси екзоцитозу. Товщина плазмолеми становить близько 10 нм (1 нм=10-9 м).
Отже, елементами клітинної оболонки є: глікокалікс, внутрішня пластинка - підмембранний або кортикальний шар цитоплазми і власне біологічна мембрана, для описання властивостей якої служить рідинно-мозаїчна модель.
Функції клітинної оболонки. До основних функцій плазмолеми слід віднести розмежування внутрішнього вмісту клітини від її мікрооточення; транспорт метаболітів, у тому числі, забезпечення асиметрії концентрації іонів натрію і калію у клітині та за її межами; примембранний метаболізм; рецепцію сигналів з боку зовнішнього середовища; забезпечення взаєморозпізнавання і взаємодії клітин з утворенням міжклітинних контактів різного ступеня складності, а також формування характерної структури клітинної поверхні.
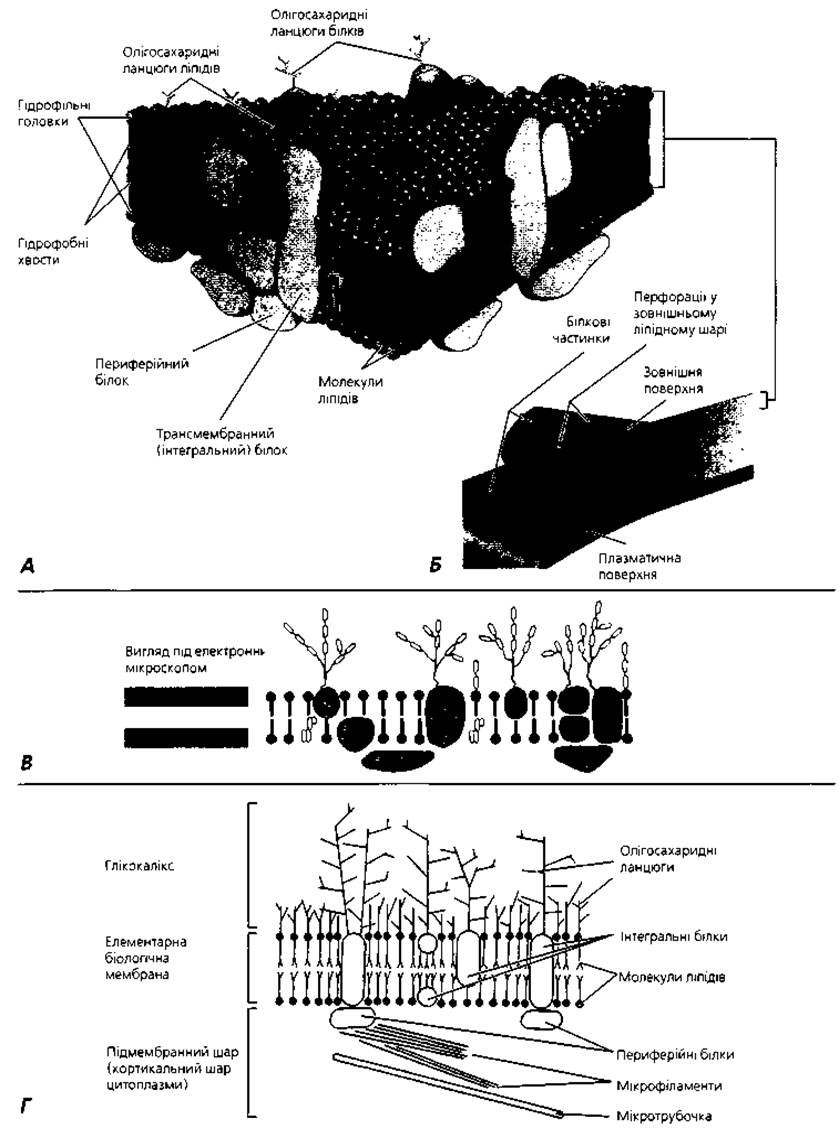
Рис.1.3. Клітинна оболонка (плазмолема): А - рідинно-мозаїчна модель Сінгер-Нікольсона; Б - розшарування плазмолеми за умови заморожування-сколювання; В - молекулярна організація плазмолеми; Г - взаємодія компонентів плазмолеми і підмембранного кортикального шару цитоплазми
Розмежування і транспорт — дві взаємопротилежні і взаємодоповнювальні функції плазмолеми. Завдяки розмежуванню із зовнішнім середовищем Клітина зберігає свою індивідуальність, завдяки транспорту речовин може жити й функціонувати. Обидва ці процеси спрямовані на підтримання постійних характеристик внутрішнього середовища (гомеостазу) клітини. Транспорт зі зовнішнього середовища всередину клітини (поглинання речовин) називається ендоцитозом, транспорт у протилежному напрямку (виведення речовин) — екзоцитозом. Невеликі молекули можуть потрапляти зі зовнішнього середовища всередину клітини або шляхом дифузії (пасивний транспорт), або за участю особливих ферментів - пермеаз плазмолеми (активний транспорт).
Великі молекули та скупчення молекул поглинаються клітиною шляхом обволікання їх певною ділянкою плазмолеми з наступним утягуванням (інтерналізацією) утвореного мішечка всередину цитоплазми. Процес поглинання таким способом твердих частинок називають фагоцитозом, поглинання частинок рідини - піноцитозом. Поглинуті частинки звичайно розщеплюються і їхні хімічні складники засвоюються клітиною. Однак можливий і такий варіант, коли частинка поглинається однією поверхнею клітини, в оточенні біомембрани проходить через цитоплазму і виводиться без змін на протилежній поверхні клітини. Таке явище має назву трансцитозу.
Виведення клітиною продуктів життєдіяльності за межі цитоплазми (екзоцитоз) поділяється на низку різновидів: секрецію, екскрецію, рекрецію, клазматоз. Секреція - це виділення клітиною продуктів її синтетичної діяльності, які необхідні для нормального функціонування органів та систем організму. Екскреція - виділення токсичних або шкідливих продуктів метаболізму, які підлягають виведенню за межі організму. Рекреція - це видалення з клітин речовин, які не змінюють своєї хімічної структури у процесі внутрішньоклітинного метаболізму (Вода, мінеральні солі). Клазматоз - видалення за межі клітини окремих її структурних компонентів.
Механізми екзоцитозу переважно мають характер прямо протилежний фаго- і піноцитозу. Наприклад, продукти синтетичної діяльності клітини нагромаджуються у вигляді оточених біомембраною скупчень (у складі мішечків і пухирців комплексу Гольджі, або пластинчастого комплексу), поступово зміщуються від центральних ділянок цитоплазми до периферії, після чого біомембрана мішечка включається у плазмолему, а вміст мішечка переходить у міжклітинний простір або зовнішнє середовище. Можливий також шлях виведення речовин у складі пухирців (без порушення цілості їх біомембрани), що має назву секреції за мерокриновим типом. Коли ж багато секреторних пухирців нагромаджуються у ділянці апікального полюса клітини і вони виділяються з відривом його, то такий спосіб секреторної діяльності має назву апокринового. У разі голокринового способу секреції Цитоплазма поступово заповнюється продуктами синтетичної діяльності й клітина перероджується в оточену плазмолемою крапельку секрету.
Примембранний метаболізм пов'язаний з наявністю на поверхні плазмолеми деяких видів клітин особливих ферментних систем, здатних розщеплювати біополімери, що контактують з ними. Такі процеси характерні, зокрема, для клітин внутрішнього вистелення тонкої кишки, на поверхні яких здійснюється так зване мембранне травлення.
Роль плазмолеми в механізмах клітинної рецепції. Сприйняття клітиною хімічних сигналів від її мікрооточення здійснюється переважно за участю спеціальних рецепторних білків плазмолеми. Для реалізації своєї дії на клітину біологічно активна речовина (наприклад гормон) повинна зв'язатися з відповідним білком плазмолеми, причому визначальна роль у забезпеченні вибірковості (специфічності) цієї взаємодії належить структурі вуглеводного компонента (олігосахаридних ланцюгів) рецепторного білка. Один із можливих шляхів подальшої передачі інформації всередину клітини пролягає через аденілатциклазну систему. Наприклад, зв'язування рецептора з відповідною хімічною речовиною зумовлює активацію розміщеного на внутрішній поверхні плазмолеми ферменту аденілатциклази, який, у свою чергу, стимулює утворення циклічного аденозинмонофосфату (цАМФ). Останній є універсальним активатором низки цитоплазматичних ферментних систем, за участю яких клітина реагує на подразнення. Слід зазначити, що складні процеси рецепції - основа взаєморозпізнавання клітин, і тому є кардинально необхідною умовою існування багатоклітинних організмів, оскільки служать командою для гальмування розмноження і руху клітин. Зауважимо, що саме втрата самоконтролю клітин за ходом росту і розмноження (відсутність контактного гальмування розмноження) - один з основних проявів злоякісного переродження.
Міжклітинні контакти. Клітинній оболонці, зокрема вуглеводним детермінантам її глікокаліксу, належить визначальна роль і в утворенні стійких контактів між клітинами (рис. 1.4). Найпростіша форма міжклітинного зв'язку має назву адгезії (прилипання, злипання). Останнім часом виявлена вирішальна роль специфічних білкових молекул - лектинів, кадгеринів і молекул клітинної адгезії, так званих САМ (від англ. Cell Adhesion Molecules) - в утворенні багатоклітинних конгломератів. Молекули лектинів, зокрема, здатні вибірково "впізнавати" вуглеводні детермінанти на поверхні сусідніх клітин і забезпечувати утворення стійких міжклітинних містків. Відстань між плазмолемами суміжних клітин у зоні простого контакту становить близько 10-20 нм.
У процесі еволюційного розвитку багатоклітинних систем значно ускладнилися форми та види міжклітинних контактів, що зумовило як зростання їхньої міцності, так і забезпечило виконання низки специфічних функцій. Один із можливих шляхів зміцнення міжклітинних контактів - збільшення площі контактних ділянок двох сусідніх клітин. У такому разі пальцеподібні вирости плазмолеми і цитоплазми однієї клітини занурюються у відповідні заглибини плазмолеми сусідньої клітини. Такий тип контакту називається пальцеподібним, зубчастим, або контактом за типом замка. Відстань між плазмолемами клітин, що контактують, така ж, як і в зоні простого контакту - 10-20 нм.
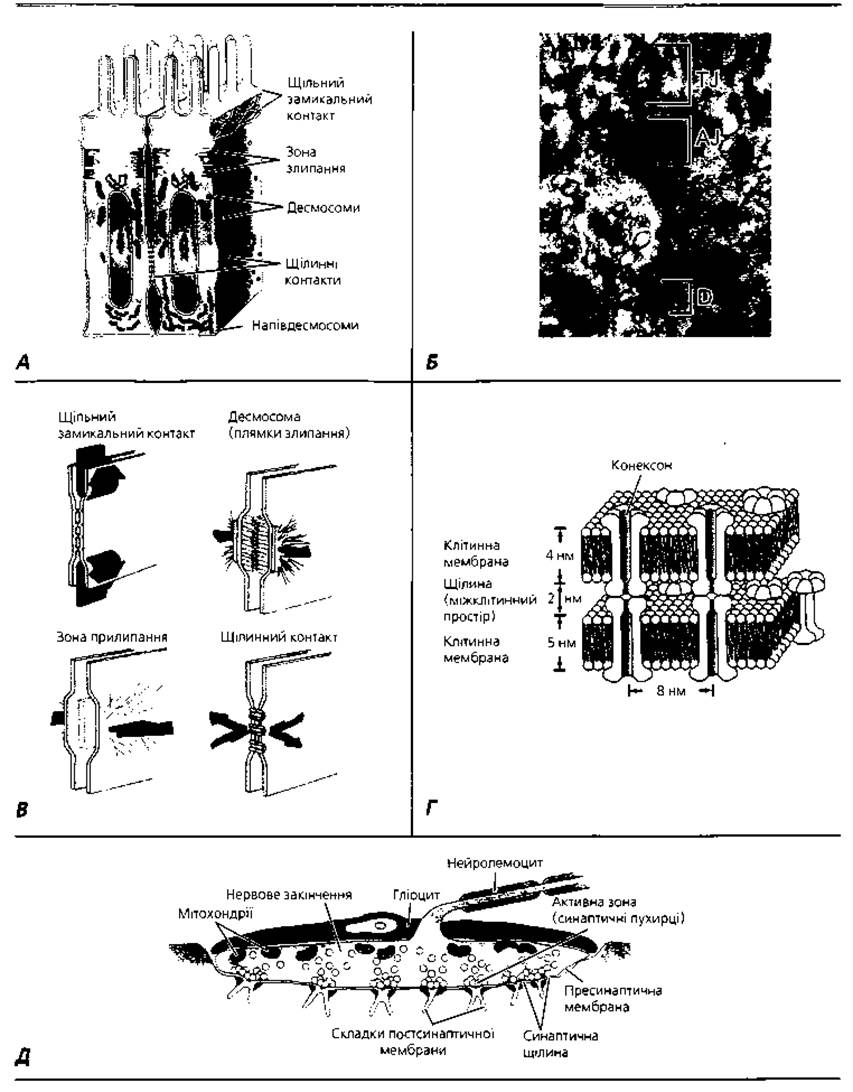
Рис. 1.4. Міжклітинні контакти: А - схема взаємодії плазмолем двох суміжних епітеліоцитів тонкої кишки; Б - електронна мікрофотографія фрагмента апікальної частини клітин, показаних на рис. 1.4,A: TJ, щільний замикальний контакт; AJ, зона прилипання; D, десмосома. X 59400; В - графічне відтворення ультраструктури та функціональних особливостей контактів різного типу; Г - молекулярна організація ділянки щілинного контакту; Д - ультраструктура синаптичного (нейром'язового) контакту, що забезпечує однобічну передачу сигналів
Подальше зміцнення зв'язку між клітинами досягається шляхом іммобілізації (знерухомлення) поверхні сусідніх ділянок плазмолеми клітин, що контактують (утворення так званих пластинок прикріплення, основу яких складає білок десмоплакін), за допомогою проміжних філаментів і кортикального шару цитоплазми. Такий тип зв'язку між клітинами має назву десмосоми і зустрічається там, де необхідно досягти максимальної міцності міжклітинних зв'язків, наприклад, у складі епітеліальної Тканини поверхні тіла. Міжклітинна щілина в ділянці десмосоми заповнена електронно-щільною речовиною, у якій розрізняють особливі трансмембранні фібрилярні структури, що складаються з білка десмоглеїну. Кінці молекул останнього приєднуються до пластинок прикріплення, за рахунок чого досягається стабілізація контакту цього типу. У ділянці десмосомних контактів ширина міжклітинної щілини становить близько 25-30 нм, діаметр десмосоми - 0,5 мкм. У місцях контакту епітеліальних клітин з базальною мембраною утворюються структури, які мають назву напівдесмосом. Якщо десмосома складається з двох, то напівдесмосома - лише з однієї пластинки прикріплення. Щілина між епітеліоцитом і базальною мембраною заповнена білками-інтегринами.
Наступна форма контакту - з утворенням щільних замикальних пластин, або щільний замикальний контакт. У ділянці такого контакту відбувається максимальне зближення плазматичних мембран сусідніх клітин. Кінці інтегральних білків плазмолем сусідніх клітин стикуються між собою, а наявний проміжок ущільнюється за рахунок іонів кальцію та фібрил, які анастомозують. Зовнішні гідрофільні шари і глікокалікс суміжних плазмолем ніби зливаються за такої умови в один суцільний шар завтовшки 2-3 нм. Щільний замикальний контакт характерний для апікальної поверхні клітин, що вистеляють травний канал. Унаслідок утворення замикальних пластин досягається повне відмежування міжклітинного простору від зовнішнього середовища. Щільні замикальні контакти спостерігаються у всіх видів епітелію (ендотелій, мезотелій, епендима, кишковий епітелій). Контакти цього типу знайдені між фібробластами, ембріональними клітинами ектодерми і мезенхіми тощо.
Базально від замикального контакту між епітеліоцитами дуже часто утворюються зони адгезії, у яких міжклітинна щілина заповнена трансмембранними білками Е-кадгеринами. Останні з'єднуються з пучками актинових філаментів, які прилягають до внутрішньоклітинної поверхні мембрани в ділянці такого контакту.
Щілинний контакт або нексус, забезпечує безпосередній обмін молекулами між сусідніми клітинами. У зонах цих контактів, які мають розміри від 0,5 до 5 мкм, гексагонально розміщені частинки - конексони з діаметром 7-8 нм і каналом шириною близько 1,5 нм у центрі. Кожний конексон складається з шести субодиниць білка конектину. Конексони вмонтовані у мембрану так, що пронизують її наскрізь. Канали двох конексонів замикаються "кінець в кінець", унаслідок чого встановлюється безпосередній хімічний зв'язок між цитоплазмами сусідніх клітин: зв'язані щілинними контактами клітини можуть вільно обмінюватися малими молекулами (неорганічними іонами, цукрами, амінокислотами, нуклеотидами, вітамінами), маса яких не перевищує 1000-1500 дальтон. У такому разі досягається своєрідна метаболічна кооперація клітин. У ділянках утворення щілинних контактів плазмолеми суміжних клітин зближені на відстань до 2-4 нм. Щілинними контактами зв'язані, зокрема, м'язові клітини міокарда, гладкі міоцити м'язової оболонки матки, овоцити і фолікулярні клітини яєчника тощо.
Синапс - спеціалізований контакт між нервовими клітинами або між нервовою клітиною і м'язом, у зоні якого відбувається передача нервового імпульсу. Основні структурні компоненти синапса: пресинаптична мембрана (ділянка плазмолеми відростка нервової клітини, з якої надходить сигнал), постсинаптична мембрана (ділянка плазмолеми клітини, яка сприймає сигнал), синаптична щілина шириною 20-30 нм (розмежовує пре- і постсинаптичну мембрани), заповнені нейромедіатором синаптичні пухирці. Функціонування синапсів забезпечує однобічну передачу інформації від клітини до клітини за допомогою медіатора (хімічного посередника).
Грунтуючись на характеристиках міжклітинних контактів, останні можна умовно поділити на три групи: адгезивні (зв'язувальні), ізолювальні та комунікаційні. До першої групи належать: простий адгезивний контакт, контакт типу замка та десмосомний контакт. Другу групу складають щільні замикальні контакти, третю - щілинний та синаптичний контакти.
Цитоплазма (Cytoplasma). Структурними компонентами цитоплазми є гіалоплазма, Органели і включення.
Гіалоплазма - найрідша частина цитоплазми, в якій містяться органели 1 включення. У загальному об'ємі цитоплазми гіалоплазма становить близько 50%. Вона включає Цитозоль (воду з розчиненими у ній неорганічними та органічними речовинами) і цитоматрикс (трабекулярну сітку волокон білкової природи товщиною 2-3 нм).
Органели - постійні структури цитоплазми, які мають певну будову і виконують спеціалізовану функцію. Органели поділяються на мікроскопічні, видимі під світловим мікроскопом, і субмікроскопічні, які можна побачити лише за допомогою електронного мікроскопа. За наявністю у складі органел біологічної мембрани їх поділяють на мембранні та немембранні. До мембранних органел належать: Мітохондрії, лізосоми, пероксисоми, ендоплазматична сітка, комплекс Гольджі. Немембранними органелами є протеасоми, Рибосоми, мікрофіламенти, мікротрубочки, центросома (клітинний центр). Ці десять органел називають органелами загального призначення, оскільки вони є в усіх видах клітин. Органели загального призначення можуть утворювати характерні конгломерати у цитоплазмі клітин. Такі конгломерати з переважним розвитком і особливою організацією органел того чи іншого виду називають спеціальними органелами (тонофібрили клітин епітелію, міофібрили м'язових клітин і волокон, нейрофібрили нервових клітин та деякі інші).
Дослідженнями останніх років установлено, що в клітині молекули білків формують мультимолекулярні комплекси характерної морфології та специфічної функції - комплексомікси - з числом індивідуальних молекул від 5 до 40 і більше. Прикладом комплексоміксів є рибосоми, апоптосоми, протеасоми тощо. Поступово вимальовується концепція, згідно з якою одні і ті ж білкові «цеглини» для виконання певної функції можуть утворювати певні комплексомікси, які після реалізації своєї активності розпадаються, а окремі білкові молекули використовуються повторно для утворення інших органел. Таким чином, внутрішньоклітинна організація усе більше нагадує мозаїку у стані постійної динамічної перебудови, в якій для складання подібних фігур у різних видах клітин використовуються різні білкові елементи. При цьому, незважаючи на індивідуальне та видове різноманіття комплектувальних білків, окремі комплексомікси і, особливо, їхні функціонально активні зони за своєю структурною організацією надзвичайно подібні навіть в еволюційно віддалених представників живого світу.
Мітохондрії - мікроскопічні мембранні органели загального призначення (рис. 1.5), основна функція яких - утворення необхідної для життєдіяльності клітини енергії та нагромадження її у складі молекул аденозинтрифосфорної кислоти (АТФ). Крім того, мітохондрії беруть участь у регуляції обміну води, депонуванні іонів кальцію, продукції попередників стероїдних гормонів. Мітохондрії відкриті німецьким дослідником Ф. Альтманом у кінці XIX ст. Під світловим мікроскопом мітохондрії мають вигляд дрібних крапочок і ниточок товщиною близько 0,5 мкм і довжиною 1-10 мкм. За допомогою електронного мікроскопа у складі кожної мітохондрії, яка має неправильну овальну або витягнуту форму, можна розрізнити дві мембрани: зовнішню гладку і внутрішню складчасту, що утворює вирости (кристи) всередину мітохондрії. Внутрішнім вмістом мітохондрії є електронно-щільна речовина, яка називається матриксом. У матриксі, а також на внутрішній мембрані мітохондрій містяться білки-Ферменти, які забезпечують синтез АТФ шляхом окисного фосфорилювання аденозиндифосфату (АДФ). Мітохондрії - єдині органели клітини, в яких знайдені молекули власної дезоксирибонуклеїнової кислоти (ДНЮ; до їх матриксу входять також різні види РНК та рибосоми.
Лізосоми - субмікроскопічні мембранні органели загального призначення (рис. 1.6), відкриті у 1955 р. Христіаном де Дювом. Основна функція лізосом - розщеплення біополімерів різного хімічного складу (так зване клітинне травлення). Для цього у лізосомах міститься набір гідролітичних ферментів (зараз їх відомо понад 60). Маркерним (визначальним) ферментом лізосом є кисла фосфатаза. Ферментні комплекси матриксу лізосоми знаходяться у замкненому мембранному мішечку діаметром близько 0,2-0,4 мкм, який перешкоджає попаданню лізосомних ферментів у гіалоплазму і запобігає самоперетравленню клітини. Залежно від ультраструктурних і функціональних особливостей лізосом їх поділяють на первинні (ферменти яких знаходяться у неактивному стані), вторинні, або фагосоми (активовані ферменти в них безпосередньо контактують з розщеплюваними біополімерами), а також залишкові тільця (оточені біомембраною нерозщеплені залишки). Слід зауважити, що лізосоми можуть брати участь як у розщепленні власних макромолекулярних комплексів клітини (таке явище має назву аутофагоцитозу), так і в перетравлюванні поглинутих клітиною частинок (гетерофагоцитоз). Недостатність того чи іншого лізосомного ферменту призводить до нагромадження у клітині аномальних біополімерів, що зумовлює розвиток, так званих, лізосомних хвороб накопичення (тезаурисмозів). До цього часу описано понад 30 різних лізосомних хвороб накопичення (табл. 2).
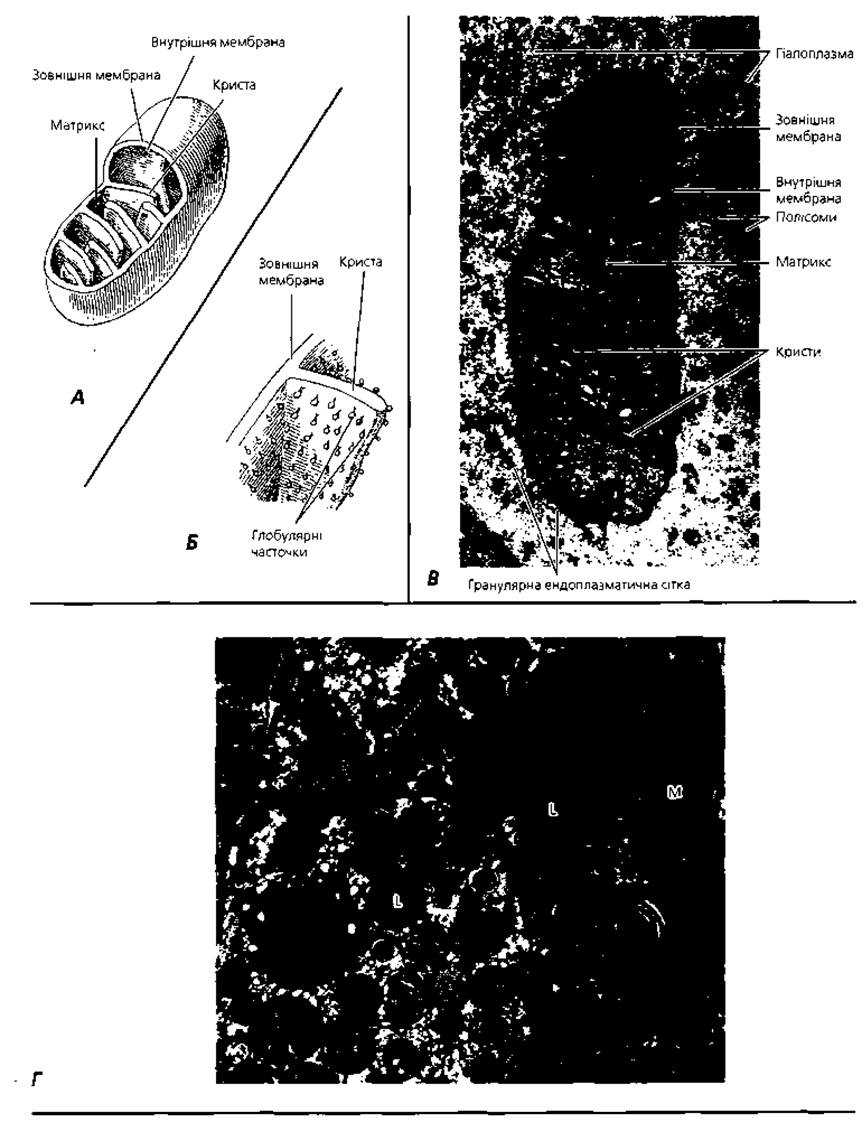
Рис. 1.5. Мітохондрії: А - об'ємна реконструкція ультраструктури; б-деталь рис. 1.5, А: схема розміщення глобулярних часточок, які забезпечують перетворення та накопичення енергії; В - електронна мікрофотографія мітохондрії епітеліоцита, х75 000; Г- крапельки ліпідів (L) та мітохондрії (М), які забезпечують їх синтез, у клітині надниркової залози, х19 000
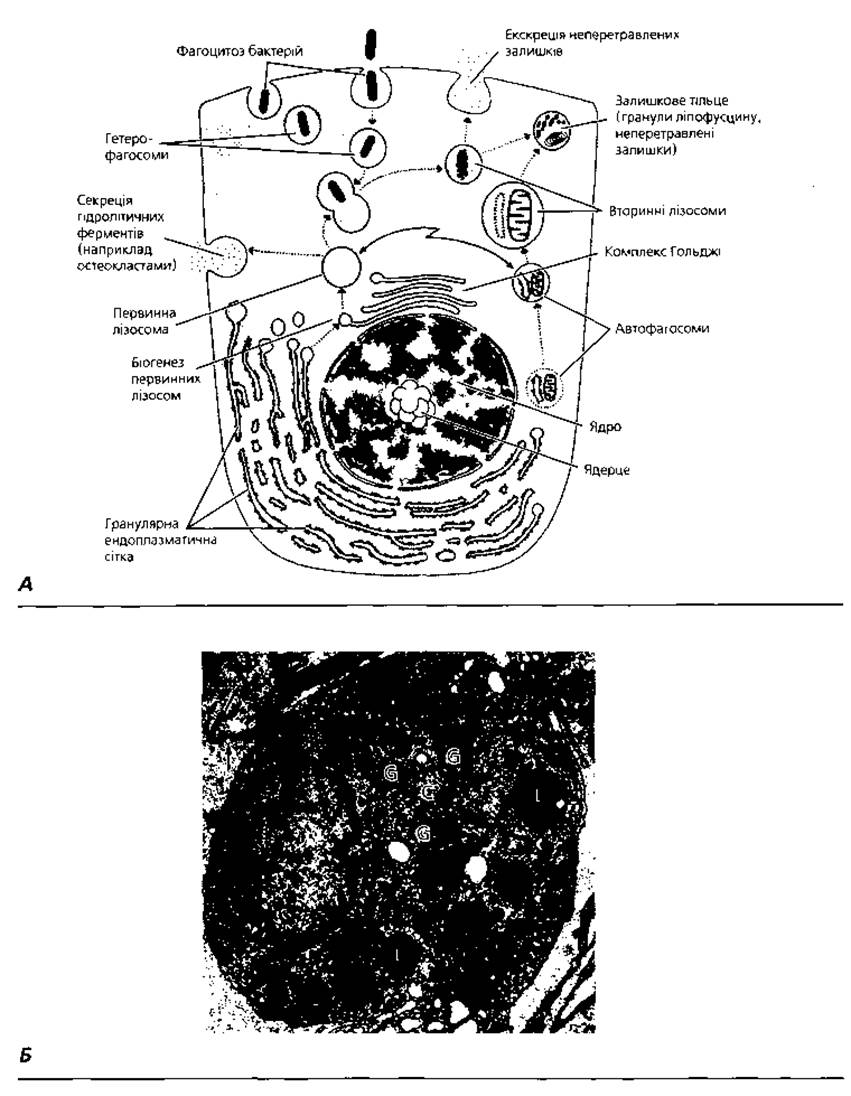
Рис. 1.6. Лізосоми: А - схема біогенезу первинних лізосом з гранулярної ендоплазматичної сітки та комплексу Гольджі, їх перетворення на вторинні лізосоми і залишкові тільця, участь у процесах секреції та екскреції; Б - електронна мікрофотографія вторинних лізосом (L) у цитоплазмі перитонеального макрофага. Стрілками показані численні вирости цитоплазми: С - центріоль; G - комплекс Гольджі, х 15 000
Таблиця 2. Приклади хвороб накопичення, пов'язаних із дефектами лізосомальних ферментів
|
Хвороба |
Відсутній фермент лізосом |
Тип клітин, які ушкоджуються |
Основні органи, які ушкоджуються |
|
Гюрлера |
a-L-ідуронідаза |
Фібробласти та остео класти накопичують дерматансульфат |
Кістки та Органи нервової системи |
|
Синдром Сан-філіппо А |
Гепаран-сульфат сульфамідаза |
Фібробласти накопичують гепаран-сульфати |
Кістки та органи нервової системи |
|
Тай-Сакса |
Гексозамінідаза А |
Нейрони накопичують гліколіпіди |
Органи нервової системи |
|
Гоше |
a -D-глікозидаза |
Макрофаги накопичують гліколіпіди |
|
|
Хвороба І-клітин |
Фосфотрансфераза |
Фібробласти та остеокласт накопичують дерматансульфат |
Кістки та органи нервової системи |
Протеасоми. У 70-ті роки XX ст. з'явилися докази того, що руйнування білків відбувається не тільки в лізосомах. Наприкінці 80-х років двома групами дослідників було показано, що білки руйнуються великими поліпротеазними комплексами, які було названо протеасомами. Зараз відомо, що кожна клітина людського тіла містить близько 30 тис. протеасом. Молекулярна маса органели - близько двох мільйонів дальтон. Кожна протеасома складається з трубкоподібної та однієї або двох регуляторних частин; останні розташовані на одному або обох кінцях органели (рис. 1.1). Трубкоподібна частина містить чотири послідовно розміщених кільця, кожне з яких складається із семи субодиниць, які оточують центральний канал протеасоми. Регуляторні частини впізнають і приєднують білки, призначені для руйнування. Вони забезпечують розкручування молекул білка та заштовхування їх у центральний канал трубкоподібної частини, де протеази розрізають їх на фрагменти різної довжини. Ці фрагменти згодом розщеплюються іншими ензимами до амінокислот, які використовуються для синтезу нових білків.
У механізмі розпізнавання білків, які повинні бути знищені у протеасомі, головну роль відіграє процес, який отримав назву убіквітинації, тобто приєднання до білкових молекул білка убіквітину. Убіквітинація складається із трьох етапів і включає три ензими: Е1, Е2 та Е3. На першому етапі Е1 активує убіквітин. На другому - за допомогою Е1 активований убіквітин приєднується до Е2. Третій етап полягає у перенесенні активованого убіквітину з Е2 на білок за допомогою Е3. Існують сотні різних ензимів Е3, кожний з яких має спорідненість з певною послідовністю амінокислотних залишків білка та робить його мішенню убіквітинації.
Численними дослідженнями показано, що убіквітинація білківта подальше їхнє руйнування у протеасомах забезпечує нормальний перебіг багатьох процесів: регуляцію внутрішньоклітинного метаболізму, імунний нагляд, звільнення від аномальних білкових молекул, Поділ клітин і міжклітинну комунікацію, розвиток і ріст організму, циркадні ритми. Порушення ж механізмів убіквітинації (наприклад, унаслідок Мутації білка Е3) та сповільнення або блокування руйнування білків у протеасомах є підгрунтям деяких спадкових аномалій (зокрема муковісцидозу), нейродегенеративних розладів (хвороба Паркінсона, хвороба Альцгеймера), багатьох вірусних захворювань, канцерогенезу.
Пероксисоми - субмікроскопічні мембранні органели загального призначення (рис. 1.7), відкриті на початку 60-х років XX ст. спільними зусиллями біохіміків і морфологів. Пероксисомам належить вирішальна роль у процесах детоксикації клітини (позбавленні її від дії токсичних продуктів обміну речовин). Утворений біомембраною мішечок округлої форми діаметром близько 0,2-0,5 мкм заповнений ферментами (матриксом), серед яких маркерним є каталаза. У центрі матриксу пероксисом за допомогою електронного мікроскопа знайдена щільна серцевина (кристалоїд), яка містить волокнисті та трубчасті макромолекулярні утвори.
Ферментні системи пероксисом спрямовані на утилізацію хімічно активного атомарного кисню (насамперед шляхом регуляції обміну і розщеплення перекису водню), а також забезпечують розщеплення етилового спирту, сечової кислоти, регуляцію обміну ліпідів.
Ендоплазматична сітка - субмікроскопічна мембранна органела загального призначення, яка утворює єдину внутрішньоцитоплазматичну циркуляційну систему, вперше описана К. Портером у 1945 р. Вона є замкненою сукупністю канальців, мішечків та цистерн, утворених суцільною (неперервною) біомембраною (рис. 1.1 і 1.8). Мембрана ендоплазматичної сітки безпосередньо контактує з плазмолемою клітини і з мембранами ядра. Розрізняють агранулярну (гладку) і гранулярну (шорстку) ендоплазматичні сітки. Агранулярна ендоплазматична сітка, діаметр канальців якої 50-100 нм, утворена лише мембраною. Гранулярна ендоплазматична сітка утворена біомембраною, до якої з боку гіалоплазми прикріплені рибосоми (рис. 1.8). Діаметр канальців гранулярної сітки від 20 до 1000 нм. Функція агранулярної ендоплазматичної сітки пов'язана з метаболізмом ліпідів і вуглеводів, детоксикацією шкідливих для клітини хімічних сполук, а також депонуванням іонів кальцію. Функція гранулярної ендоплазматичної сітки зумовлена наявністю рибосом і полягає у біосинтезі білків як для потреб самої клітини, так і для виведення за її межі. Крім виконання метаболічної та циркуляторної функції ендоплазматична сітка - єдина органела, в якій відбувається новотворення мембранних структур клітини. Синтезовані гранулярною ендоплазматичною сіткою компоненти біомембрани можуть включатися до складу мішечків лізосом, пероксисом, елементів комплексу Гольджі, плазмолеми, оболонки ядра, а також використовуватися для самовідтворення елементів ендоплазматичної сітки.
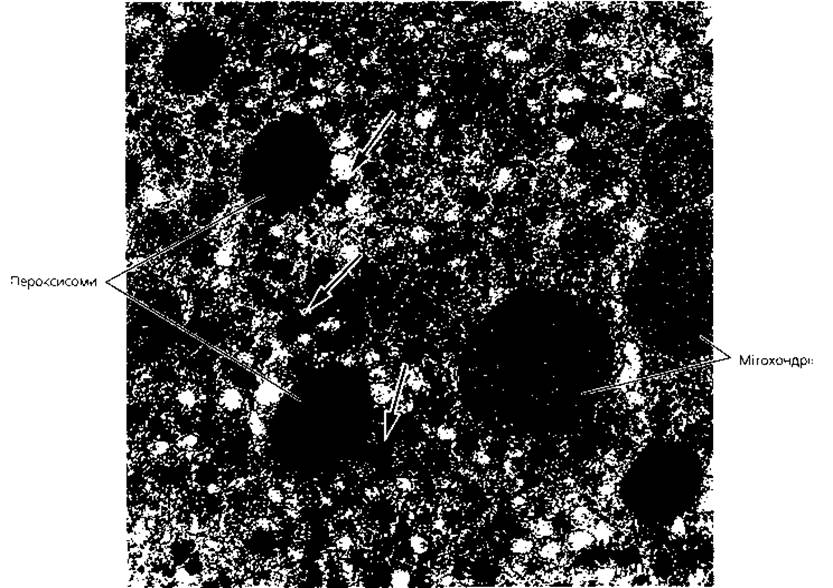
Рис. 1.7. Пероксисоми. Електронна мікрофотографія пероксисом та мітохондрій у цитоплазмі гепатоцита. Стрілками показано включення глікогену, х30 000.
Комплекс Гольджі або пластинчастий комплекс - мікроскопічна мембранна органела загального призначення (рис. 1.9), в якій завершується процес формування продуктів синтетичної діяльності клітини, зокрема, здійснюється їх кінцеве глікозування. Комплекс Гольджі нагромаджує секреторні речовини і забезпечує їх виведення за межі клітини. Органела була названа іменем італійського гістолога Камілло Гольджі, який у 1898 р. описав цей комплекс у складі нервових клітин. Морфологічно пластинчастий комплекс - це сукупність пов'язаних між собою цистерн товщиною близько 25 нм, сплющених у центральній частині й розширених на периферії. Проміжки між окремими цистернами становлять 20-25 нм. Від розширених країв цистерн відокремлюються дрібні пухирці (рис. 1.6). Окрема сукупність таких цистерн і пухирців має назву диктіосоми. Образно диктіосому порівнюють зі стосом тарілок, повернених опуклим боком до ядра. В одній клітині може бути кілька диктіосом, відокремлених одна від одної прошарками гіалоплазми. Завершується синтез (дозрівання) секреторних продуктів під час переміщення їх із цистерн, розташованих поблизу ядра (так звана цис-локалізація цистерн), у напрямку плазмолеми (транс-локалізація цистерн). Пухирці, які відокремлюються від країв цистерн, містять сформовані, готові до виведення із клітини секреторні продукти. Нагромаджені у складі пухирців макромолекулярні комплекси виводяться шляхом умонтовування мембран пухирців у плазмолему або виштовхуванням сформованих (зрілих) пухирців за межі клітини.
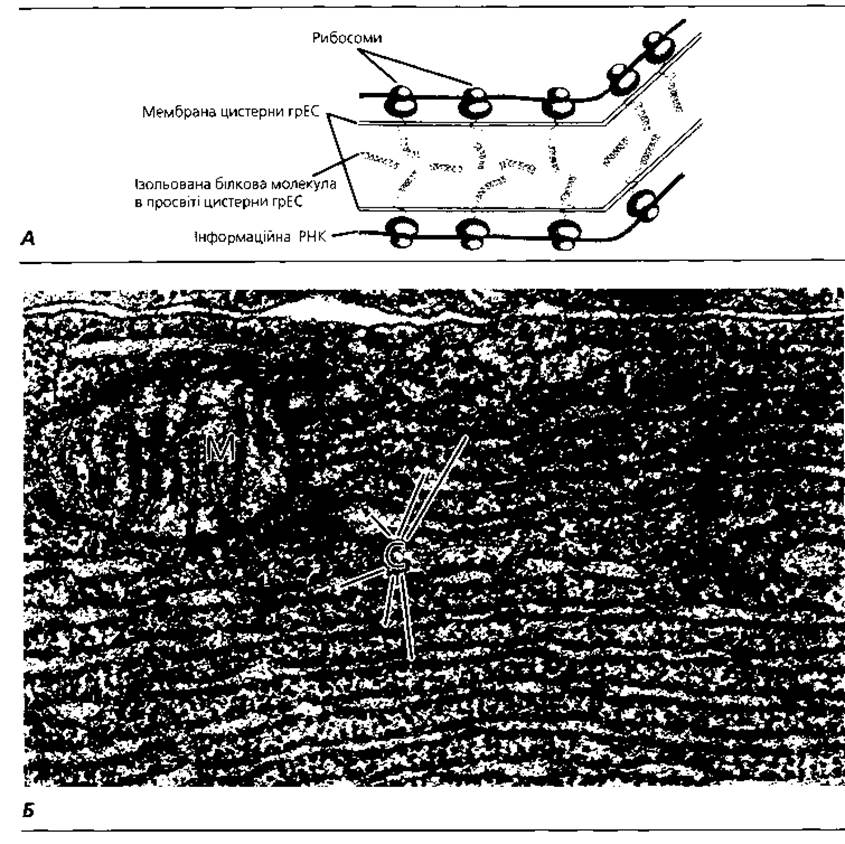
Рис. 1.8. Гранулярна ендоплазматична сітка (грЕС): А - схема біосинтезу і накопичення білків у цистерні грЕС; Б - електронна мікрофотографія грЕС у головній клітині залози шлунка: С - цистерни грЕС; М - мітохондрія
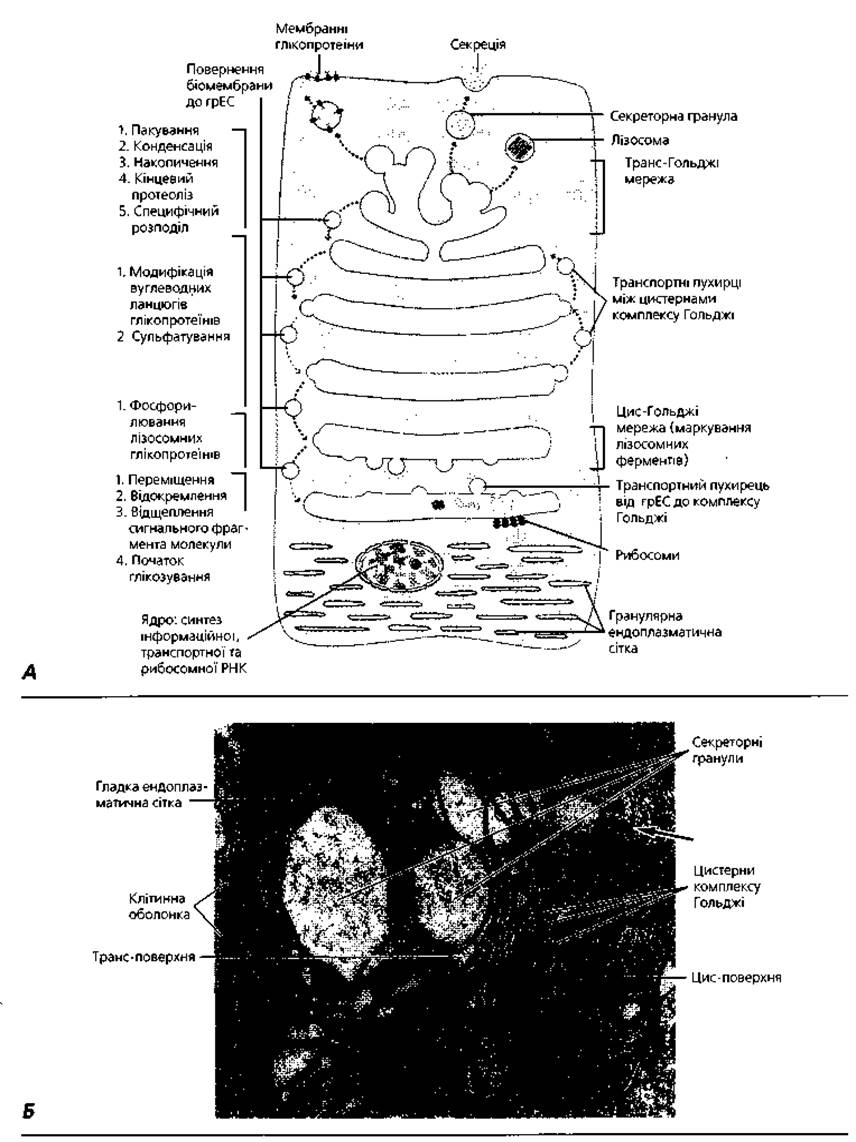
Рис. 1.9. Комплекс Гольджі: А - схема взаємозв'язку елементів комплексу Гольджі з гранулярною ендоплазматичною сіткою, лізосомами, секреторними гранулами та пухирцями; Б - електронна мікрофотографія пластинчастого комплексу слизопродукувальної клітини. Стрілкою показана цистерна гранулярної ендоплазматичної сітки, х30 000.
Рибосоми - субмікроскопічні немембранні органели загального призначення (рис. 1.10), у яких амінокислоти сполучаються, утворюючи пептидний ланцюг, тобто синтезуються білкові молекули. Уперше рибосоми у складі гранулярної ендоплазматичної сітки описав Дж. Паладе. Морфологічно у рибосомах налічуються дві субодиниці. Їх сполучення утворює структуру, яка за формою нагадує гриб. Діаметр рибосом - близько 20 нм. З хімічної точки зору рибосома - це рибонуклеопротеїновий комплекс рибосомної РНК і білка у співвідношенні 1:1. У разі дії ушкоджувальних чинників або порушення електролітного гомеостазу клітини (брак іонів магнію) спостерігається розпад рибосоми на субодиниці (дезагрегація), біологічна активність її при цьому втрачається. Кілька рибосом, "нанизаних" на спільну нитку інформаційної РНК, називають полісомами. На завислих у гіалоплазмі полісомах здійснюється Синтез білків переважно для внутрішніх потреб клітини. Полісоми, пов'язані з мембранами ендоплазматичної сітки, переважно синтезують білки для виведення за межі клітини.
Мікрофіламенти - субмікроскопічні немембранні органели загального призначення (рис. 1.11), які виконують роль цитоскелета, а також скоротливого апарату клітини. Мікрофіламенти - це тонкі волоконця, побудовані з білків актину (тонкі філаменти, діаметр близько 5 нм), міозину (товсті філаменти, діаметр близько 25 нм), тропоміозину або альфа-актиніну. Вони розміщені переважно в кортикальній (підмембранній) зоні клітини та у складі її цитоплазматичних виростів. їхня функція - скоротливо-рухова. Так звані проміжні мікрофіламенти мають діаметр 10-15 нм. Білок, з якого побудовані проміжні філаменти, є суто специфічною ознакою клітин того чи іншого типу. Наприклад, кератин служить гістохімічним маркером клітин епітелію, віментин - сполучної тканини, десмін - м'язів, білок нейрофіламентів і гліальний фібрилярний білок - нервової тканини. Проміжні мікрофіламенти в основному відповідають за збереження клітиною своєї форми. В останні роки підтверджений їх зв'язок з регуляцією активності геному і процесами клітинної диференціації (табл. 3). У складі спеціалізованих клітин мікрофіламенти можуть утворювати своєрідні пучки складнішої будови (тонофібрили епітеліальних клітин, міофібрили м'язів, нейрофібрили нервових клітин). Унаслідок особливої організації тонофібрили, міофібрили та нейрофібрили належать до спеціальних органел відповідних видів клітин.
Мікротрубочки - субмікроскопічні немембранні органели загального призначення (рис. 1.12), основна функція яких полягає у забезпеченні рухомості клітинних органел, а також в утворенні цитоскелета (рис. 1.1 ). Мікротрубочки побудовані з глобулярних білків тубулінів, молекули яких здатні до полімеризації. Особливим способом "нанизуючись" одна на одну, окремі молекули утворюють своєрідні "намистинки". 13 ниток "намистинок", які розміщені паралельно, формують порожнистий циліндр діаметром близько 25 нм з внутрішнім просвітом 15 нм. Товщина стінки циліндра відповідає діаметру однієї молекули тубуліну і становить 5 нм. Полімеризація молекул тубулінів є динамічним процесом, який припиняється під дією несприятливих чинників зовнішнього середовища (зниження температури, обробка колхіцином). Часткова деполімеризація мікротрубочок призводить до їх вкорочення, повна - до розпаду (ДИСОЦІАЦІЇ) на окремі молекули тубулінів. Мікротрубочки є основою будови центросоми, а також таких спеціалізованих структур, як війки та джгутики.
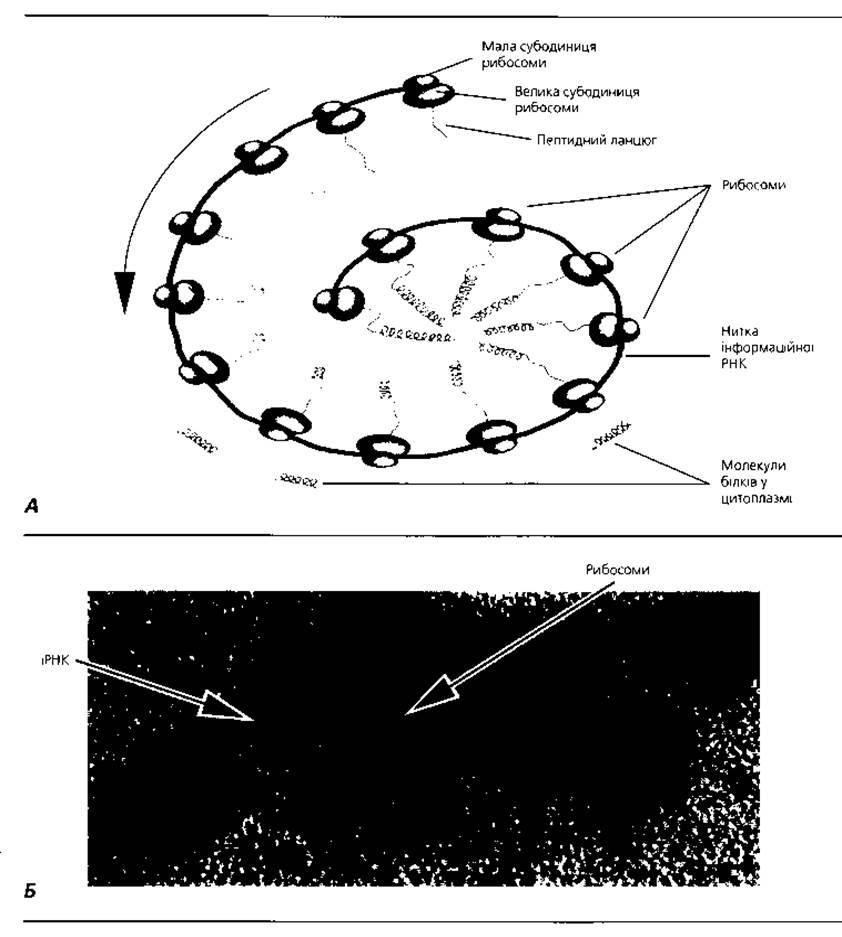
Рис. 1.10. Рибосоми: А - схематичне відтворення організації вільних рибосом (полісоми), які забезпечують біосинтез цитоплазматичних білків; Б - електронна мікрофотографія полісоми, яка включає п'ять рибосом, приєднаних до нитки інформаційної РНК, що синтезує глобіновий ланцюг гемоглобіну, х 400 000
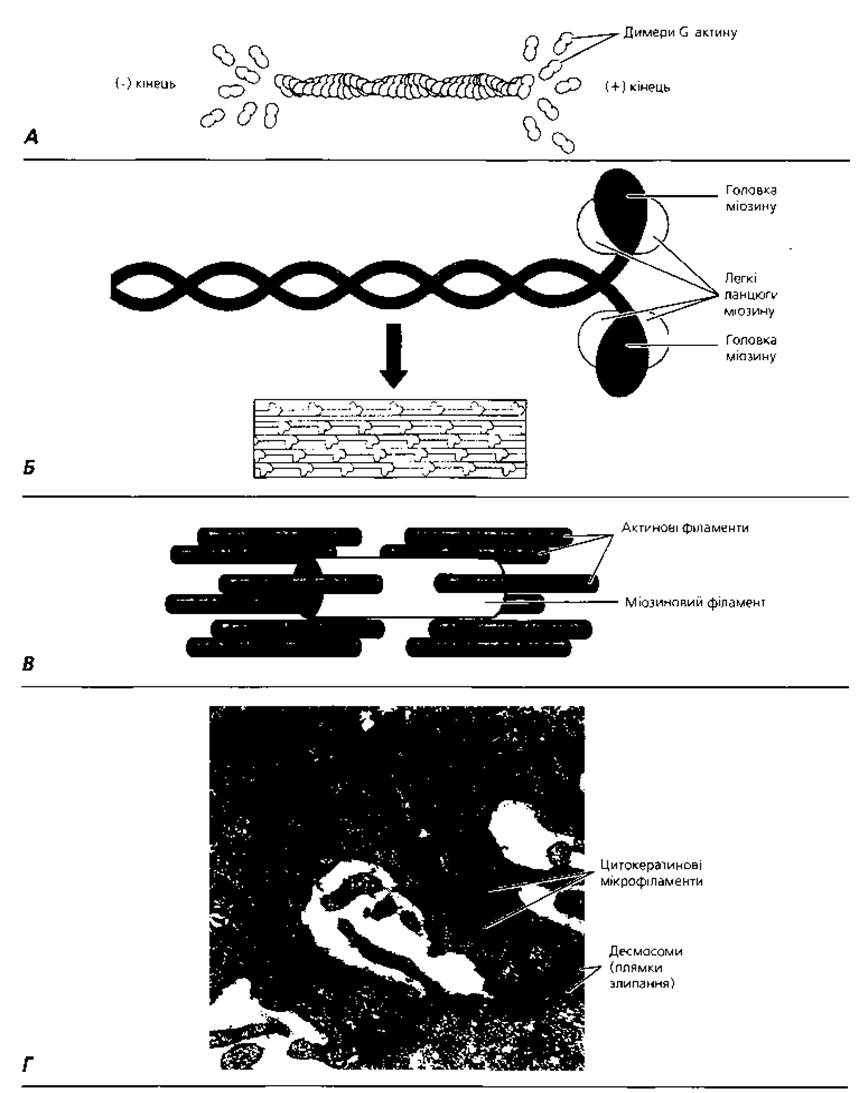
Рис. 1.11. Мікрофіламенти: А - актиновий (тонкий) філамент: приєднання димерів G-актину на (+) кінці і дисоціація на (-) кінці забезпечує зміну довжини філамента у залежності від потреб клітини; Б - самозбирання міозинового (товстого) філамента з молекул міозину; В - співвідношення актинових і міозинового філаментів у скороченому м'язовому волокні; Г - електронна мікрофотографія цитокератинових (проміжних) філаментів ділянки десмосомних контактів між клітинами

Рис. 1.12. Мікротрубочки: А- схема самозбирання на (+) кінці та дисоціації на (-) кінці мікротрубочки внаслідок приєднання або відщеплення димерів а- і ß-тубуліну, що забезпечує видовження, вкорочення або саморозпад мікротрубочки залежно від потреб клітини; Б - електронна мікрофотографія поперечно зрізаних мікротрубочок: 13 тубулінових димерів укладені по спіралі; В - електронна мікрофотографія поздовжньо орієнтованих мікрофіламентів (MF) та мікротрубочок (МТ) у цитоплазмі фібробласта,х60 000
Таблиця 3. Перелік найпоширеніших антигенів, що використовуються для імуноцитохімічної діагностики та подальшого Лікування
|
Антигени |
Вид патології |
|
Цитокератини |
Недиференційовані пухлини епітеліального походження, карциноми, аденокарциноми |
|
Гліальні фібрилярні кислі білки |
Гліальні пухлини |
|
Віментин |
Пухлини сполучної тканини |
|
Десмін |
Пухлини м'язової тканини |
|
Пептидні гормони |
Пухлини, що продукують білкові або пептидні гормони |
|
Карциноембріональний антиген |
Залозисті пухлини, переважно травного каналу та повітроносних шляхів |
|
Простато-специфічний антиген |
Пухлини передміхурової залози |
|
Рецептори до стероїдних гормонів |
Пухлини повітроносних шляхів |
|
Антигени вірусів |
Вірусні інфекції |
Центросома (клітинний центр) - мікроскопічна немембранна органела загального призначення (рис. 1.13), описана В. Флемінгом у 1875 р., яка забезпечує розходження хромосом під час поділу клітини. У клітині, що не готується до поділу, центросома розміщена поблизу ядра і складається із двох повністю сформованих центріолей, оточених центросферою. Дві розташовані поряд центріолі називають диплосомою. Кожна центріоль у своїй основі містить дев'ять триплетів паралельно орієнтованих мікротрубочок, які у просторовому зображенні формують циліндр діаметром 200 нм і довжиною близько 500 нм (рис. 1.1). Крім мікротрубочок до складу центріолі входять специфічні макромолекулярні утвори, так звані ручки, за допомогою яких триплети пов'язані між собою. У складі "ручок" міститься білок динеїн, який має АТФ-азну активність і якому належить вирішальна роль у механізмах реалізації рухових функцій центріолей. Довгі осі обох центріолей розташовані у взаємно перпендикулярних площинах. Центросфера являє собою позбавлену органел гіалоплазму навколо центріолей, яку в радіальному напрямку пронизують мікрофіламенти і мікротрубочки. Під час підготовки клітини до поділу відбувається подвоєння (дуплікація) центріолей з наступним розходженням кожної новоутвореної пари до полюсів клітини. Основою функціональної активності центросоми є механізм стимулювання полімеризації тубулінів, що зумовлює ріст наявних та утворення нових мікротрубочок.

Рис. 1.13. Центросома: А - схема самоорганізації мікротрубочок (9x3) у центріоль та клітинний центр - центросому (масштаб не збережено); Б - електронна мікрофотографія поперечно зрізаної центріолі фібробласта: mt, мікротрубочки; рс, парацентріолярний матеріал; В - електронна мікрофотографія сперматоцита в метафазі мітозу: від пари центріолей на кожному з полюсів клітини до екваторіальної пластинки хромосом тягнуться мікротрубочки. Стрілками показано ділянки приєднання мікротрубочок до центромерних ділянок хромосом, х19 000
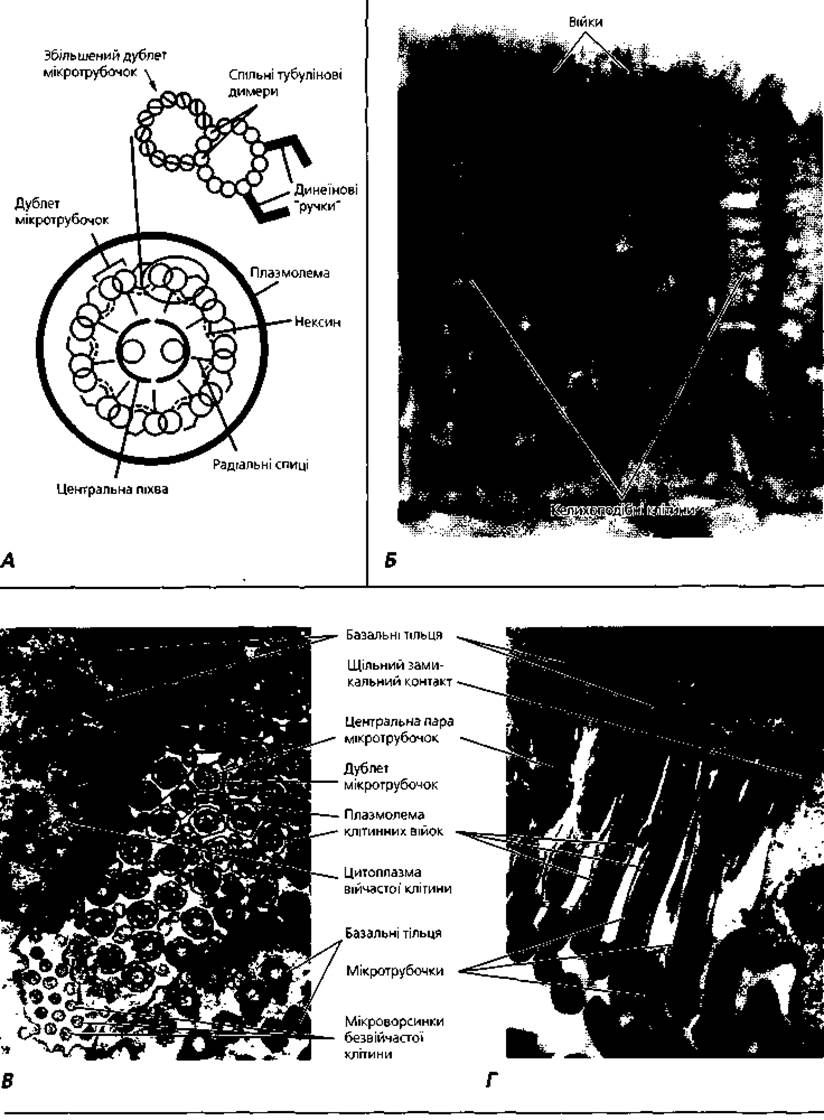
Рис. 1.14. Війки: А - схема організації мікротрубонок аксонеми (9x2+2) клітинної війки; Б-світлова мікроскопія респіраторного війчастого епітелію трахеї, х1200. Електронні мікрофотографії поперечно (Б) та поздовжньо (Г) зрізаних війок епітеліоцитів маткової труби самки, х25 000
Війки (рис. 1.14) і джгутики — це тонкі вирости цитоплазми, основою яких є високоорганізована система мікротрубочок. Довжина війок становить 5-10 мкм, джгутиків - досягає 150 мкм. Діаметр обох цих структур приблизно однаковий і становить близько 200 нм. Ззовні війку покриває плазмолема, всередині її проходить осьова нитка - аксонема. Аксонема містить дев'ять пар (дублетів) мікротрубочок, які формують порожнистий циліндр діаметром близько 150 нм. У центрі цього циліндра локалізується центральна пара мікротрубочок. Систему мікротрубочок аксонеми описують формулою (9х2)+2, Біля основи війки в ділянці її переходу в цитоплазму розташоване, так зване, базальне тільце, яке за будовою нагадує центріоль і складається з дев'яти триплетів мікротрубочок. Формула системи мікротрубочок базального тільця має вигляд (9хЗ)+0, тобто така сама, як у центріолі. Часто біля основи війки знаходяться два базальні тільця, довгі осі яких розміщені під прямим кутом одна до одної. Базальне тільце й аксонема пов'язані між собою: дві мікротрубочки з кожного триплету базального тільця продовжуються у дублет мікротрубочок аксонеми. Незначні зміщення дублетів мікротрубочок можуть зумовити вигинання усієї війки або джгутика. У різних клітинах рух війок і джгутиків може нагадувати рух маятника, бути лійкоподібним, хвилеподібним тощо. Вільні клітини, що мають війки і джгутики, можуть переміщатися у просторі (наприклад сперматозоїди). Фіксовані клітини з війками на апікальній поверхні рухом своїх війок можуть транспортувати рідину, слиз або завислі у них частинки чи клітини (наприклад, війчасті клітини дихальних шляхів, маткових труб тощо).
Таблиця 4. Деякі хвороби людини і тварин, спричинені ушкодженням цитоплазматичних компонентів
|
Ушкоджений клітинний компонент |
Хвороба |
Молекулярний дефект |
Морфологічні зміни |
Характерні симптоми |
|
Мітохондрії |
Мітохондріальна цитопатія |
Ушкодження окисного фосфорилювання |
Збільшення розмірів і кількості мітохондрій у клітинах м'язової тканини та їх похідних |
Високий рівень основного обміну без гіпертироїдизму |
|
Мікротрубочки |
Синдром Картагенера |
Відсутність динеїну в джгутиках і війках |
Відсутність ручок у дублетах мікротрубочок |
Нерухомість джгутиків і війок, що призводить до чоловічої стерильності та хронічних респіраторних інфекцій |
|
Мишиний діабет |
Зменшення кількості тубуліну у ß-клітинах острівців підшлункової залози |
Зменшення мікротрубочок у ß-клітинах острівців підшлункової залози |
Діабет |
|
|
Лізосоми |
Метахроматична лейкодистрофія |
Відсутність лізосомної фосфатази |
Накопичення ліпідів (цереброзидів) у тканинах |
Зниження рухової активності та Інтелекту |
|
Хвороба Гюрлера |
Відсутність лізосомальної a-L-ідуронідази |
Накопичення дерматан-сульфатів у фібробластах та остеобластах |
Затримка росту та розумового розвитку |
|
|
Комплекс Гольджі |
Хвороба І -клітин |
Недостатність фосфотрансферази |
Накопичення дерматан-сульфатів у фібробластах та остеобластах |
Дефекти кісток, затримка психомоторного розвитку |
|
Секреторні гранули |
Проінсуліновий діабет |
Дефект ферменту, що розщеплює проінсулін |
Немає |
Діабет |
Ушкодження тих або інших органел клітин призводить до різних захворювань (табл. 4).
Включення, на відміну від органел, не є постійними структурними компонентами цитоплазми і не мають чітко визначеної будови. Включення бувають екзо- та ендогенні. Останні залежно від їхнього функціонального призначення поділяють на екскреторні, трофічні, пігментні тощо. Власне кажучи, включення можна розглядати як макромолекулярні конгломерати, які клітина у тих чи інших умовах свого функціонування нагромаджує у цитоплазмі.
Терміни для запам'ятовування
1. Плазмолема. 2. Глікокалікс. 3. Кортикальний (підмембранний) шар. 4. Ендоцитоз. 5. Фагоцитоз. 6. Піноцитоз. 7. Трансцитоз. 8. Секреція. 9. Мерокриновий тип секреції. 10. Апокриновий тип секреції. 11. Голокриновий тип секреції. 12. Екскреція. 13. Рекреція. 14. Клазматоз. 15. Лектин. 16. Кадгерин. 17. Адгезія. 18. Десмосома. 19. Нексус. 20. Конексон. 21. Конектин. 22. Синапс. 23. Гіалоплазма. 24. Цитозоль.25. Цитоматрикс. 26. Органела. 27. Мітохондрія. 28. Лізосома. 29. Фагосома. 30. Залишкове тільце. 31. Ауто- фагоцитоз. 32. Гетерофагоцитоз. 33. Протеасома. 34. Пероксисома. 35. Ендоплазматична сітка. 36. Пластинчастий комплекс. 37. Диктіосома. 38. Рибосома. 38. Полісома. 40. Мікрофіламент. 41. Тонофібрила. 42. Міофібрила. 43. Нейрофібрила. 44. Кератин. 45. Віментин. 46. Десмін. 47. Мікротрубочка. 48. Тубулін. 49. Центросома (клітинний центр). 50. Центріоль. 51. Диплосома. 52. Центросфера. 53. Війка. 54. Аксонема. 55. Базальне тільце. 56. Джгутик. 57. Включення.
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.