Гістологія людини - О.Д. Луцик 2003
Загальна гістологія
Нервова тканина
Нервова тканина (textus nervosus) належить до спеціальних тканин, її елементи здатні сприймати подразнення, трансформувати це подразнення у нервовий імпульс, швидко його передавати, зберігати інформацію, продукувати БІОЛОГІЧНО АКТИВНІ РЕЧОВИНИ, завдяки чому нервова тканина забезпечує узгоджену діяльність органів і систем організму та його адаптацію до умов зовнішнього середовища. Нервова тканина побудована з нервових клітин (нейронів, нейроцитів) та допоміжних елементів, які об'єднуються під назвою нейроглії (рис. 3.49).
Нейрони (рис. 3.49,3.50,3.51) є морфологічними і функціональними одиницями нервової тканини. Складаються з тіла (перикаріону) і відростків. Наявність останніх є найхарактернішою ознакою нервових клітин. Саме відростки забезпечують проведення нервового імпульсу часто на досить значну відстань, тому довжина їх коливається від кількох мікрометрів до 1-1,5 м. Нейрони звичайно не здатні до мітотичного поділу, мають довгий життєвий цикл. Термін їхнього життя збігається із терміном життя індивіду. Але це стосується не всіх клітин. Нещодавно стало відомо, що в дорослих ссавців, із людиною включно, утворюються нові Нервові клітини. Але нейрогенез (утворення нейронів зі стовбурових клітин) відбувається лише в двох ділянках: субвентрикулярній зоні мозку та субгранулярній зоні гіпокампа. На певних етапах онтогенезу (під час становлення органів центральної нервової системи) відбувається також масова запрограмована (фізіологічна) загибель нейронів. Після народження у людини в середньому за рік гине близько 10 млн клітин, а протягом життя мозок втрачає приблизно 0,1% усіх нейронів (загальна кількість нейронів у людини не менше ніж 1012 (трильйон)). Розміри перикаріона нейронів дуже різноманітні - від 5-8 (клітини-зерна мозочка) до 120 мкм (гігантські пірамідні нейрони кори великого мозку). Серед відростків нервових клітин розрізняють аксони і дендрити.
Аксон (нейрит, неврит) - це довгий відросток, довжина якого може сягати 1,5 м. Назва його походить від грецького "аксіс" - вісь. Він завжди у клітині лише один. Діаметр аксона на всій довжині є незмінним, він не розгалужується, але може давати колатералі, що мають інший напрямок. Починається аксон від перикаріона так званим аксонним горбиком - конічною ділянкою цитоплазми, яка не містить хроматофільної субстанції (див. нижче). Закінчується аксон термінальним розгалуженням, що містить синаптичні пухирці. Аксон проводить нервовий імпульсу напрямку від тіла клітини. Експресія нейромодуліну (GAP-43) - специфічного для аксона фосфобілка - є ознакою початку диференціації нейронів. Спочатку утворюються короткі відростки, які потенційно можуть стати або аксоном, або дендритами. Відросток, що нагромаджує білок GAP-43, у подальшому стає аксоном. Об'єм аксона може сягати 99% усього об'єма нейрона.

Рис. 3.49. Клітинні елементи нервової тканини: А - нейрони (нейроцити); Б - клітини нейроглії. Співвідношення розмірів клітин не дотримано

Рис. 3.50. А - схема структурної організації мультиполярного мотонейрона; Б - напівсхематичне відтворення гістологічного препарату нервової тканини: мультиполярні мотонейрони переднього рогу спинного мозку, х 400

Рис. 3.51. Світлова мікроскопія нейроцитів: А - ланцюг нейронів сполучнотканинної строми ворсинки тонкої кишки, імпрегнація сріблом, х 100; Б - нейрони парасимпатичного ганглію підщелепної слинної залози, гістохімічна реакція з конканаваліном А, х 400; В - мотонейрон переднього рогу спинного мозку, х 1200
Дендрити - це здебільшого короткі відростки (хоча зустрічаються і довгі дендрити), які деревоподібно розгалужуються. Назва їх походить від грецького слова "дендрон" - дерево. Основи дендритів мають конічне розширення. Ці відростки передають нервовий імпульс у напрямку до тіла клітини.
Нервові клітини містять у центрі перикаріона одне велике округле світле Ядро з малою кількістю гетерохроматину, одним або кількома ядерцями. Разом з тим у нейронах деяких гангліїв вегетативної нервової системи налічується до 15 ядер. Цитоплазма нервової клітини (нейроплазма) містить три типи організованих структур: загальні Органели, включення та спеціальні органели. Включеннями нейроплазми можуть бути Вуглеводи (глікоген), пігментні речовини (ліпофусцин, меланін) та різноманітні секреторні продукти (у нейросекреторних клітинах). Спеціальними органелами нейронів є хроматофільна субстанція і нейрофібрили.
Хроматофільна субстанція під світловим мікроскопом має вигляд різних за розмірами і формою грудочок та зерен, які фарбуються базофільно, локалізовані у перикаріоні та дендритах нейронів і ніколи не виявляються в аксонах та їхніх початкових сегментах. Хроматофільну субстанцію уперше описав Ф. Ніссль 1889 р., у зв'язку з чим вона була названа його іменем (речовина Ніссля). Й. Леношек (1895) дав їй назву «тигроїд». Хроматофільну субстанцію також називають базофільною речовиною. Під електронним мікроскопом ця структура виявляється гранулярною ендоплазматичною сіткою з паралельним розташуванням її сплощених цистерн (так звана ергастоплазма), де інтенсивно синтезується білок, необхідний для функціонування нервової клітини. Хроматофільна субстанція є показником функціонального стану нейрона. Вона може зникати у разі виснаження нервової клітини (так званий хроматоліз, або тигроліз), а потім відновлюватися.
В аксонах, що не містять органел білкового синтезу, цитоплазма постійно переміщується від перикаріона до терміналей із швидкістю 1-3 мм на добу. Це так званий повільний аксонний транспорт, за рахунок якого відбувається транспортування білків, наприклад ферментів, необхідних для синтезу медіаторів у синаптичних закінченнях. Крім того, існує швидкий аксонний транспорт (5-10 мм/год), що переносить переважно речовини, необхідні для реалізації синаптичної функції, дендритний транспорт (швидкість 3 мм/год) і ретроградний потік, за допомогою якого низка компонентів цитоплазми повертається із закінчень у тіло клітини. Транспорт речовин по відростках нейронів забезпечують головним чином мікротрубочки.
У перикаріоні і дендритах мікротрубочки не мають спрямованої орієнтації, в той час як в аксоні вони (+)-кінцем обернені до терміналей, а (-)-кінцем — до перикаріона. Характер орієнтації мікротрубочок має важливе значення для розподілу різних органел у відростках нейрона. До (+)-кінця рухаються Мітохондрії і секреторні пухирці, а до (-)-кінця — Рибосоми, мультивезикулярні тільця, елементи комплексу Гольджі. Стабілізацію і паралельне розташування мікротрубочок забезпечує білок Тау. Його надмірне фосфорилювання призводить до дезорієнтації нейротрубочок і є однією з причин розвитку хвороби Альцгаймера.
Нейрофібрили можна виявити у цитоплазмі у випадку імпрегнації солями срібла. Вони мають вигляд тонких ниток діаметром 0,3-0,5 мкм, утворюють щільну сітку в перикаріоні і мають паралельну орієнтацію у складі дендритів та нейритів, включаючи їх найтонші кінцеві розгалуження. Методом електронної мікроскопії виявлено, що нейрофібрилам відповідають пучки нейрофіламентів (проміжних мікрофіламентів) діаметром 6—10 нм і нейротубули (мікротрубочки) діаметром 24 нм. Мікрофіламенти і мікротрубочки належать до системи цитоскелета нейронів. У складі останнього міститься також білок спектрин, який є аналогом спектрину еритроцитів.
Морфологічна Класифікація нейронів грунтується на кількості відростків. За цією ознакою нервові клітини поділяють на такі різновиди:
1) уніполярні (мають єдиний відросток, який є аксоном);
2) біполярні (мають два відростки - аксон і дендрит);
3) псевдоуніполярні (мають один відросток, який на певній відстані від тіла клітини поділяється на аксон і дендрит, так що фактично Клітина має два відростки, як і біполярна);
4) мультиполярні (мають багато відростків, один з яких є аксоном, а всі інші дендритами).
В організмі людини переважна більшість нейронів є мультиполярними; біполярні клітини є лише у сітківці ока і в гангліях присінково-завиткового нерва, а псевдоуніполярні - у спинномозкових вузлах. Уніполярні клітини в тілі людини не виявлені. Один відросток мають лише нейробласти - малодиференційовані нервові клітини.
Функціональна класифікація нейронів грунтується на функції нервової клітини у складі рефлекторної дуги. Згідно з цією класифікацією розрізняють такі види нейронів:
1) аферентні (рецепторні, чутливі) сприймають подразнення і трансформують його у нервовий імпульс;
2) асоціативні (вставні) передають нервовий імпульс між нейронами;
3) еферентні (моторні, рухові, секреторні) забезпечують передачу нервового імпульсу на робочу структуру.
Рефлекторна дуга - це ланцюжок нервових клітин, який передає нервовий імпульс від чутливого нервового закінчення (рецептора) до рухового нервового закінчення (ефектора), що розташоване у робочому органі. Найпростіша рефлекторна дуга складається із двох нейронів: аферентного, дендрит якого закінчується рецептором, а аксон передає імпульс на дендрит еферентного нейрона; еферентного, який своїм аксоном передає імпульс до ефектора у робочому органі. Складні рефлекторні дуги мають між аферентним і еферентним нейронами одну або кілька асоціативних нервових клітин. Нервове збудження по рефлекторній дузі передається лише в одному напрямку, що має назву фізіологічної (або динамічної) поляризації нейронів. Ізольований нейрон, як показав О.І. Бабухін, здатний проводити імпульс у будь-якому напрямку. Однонаправленість передачі імпульсу в межах рефлекторної дуги зумовлена структурою міжнейронного контакту, що має назву синапса.
Синапси. У складі синапса (рис. З.Б2, 3.53) є дві частини - пресинаптична та постсинаптична, між якими розташована синаптична щілина. Преси- наптична частина (або полюс) утворена термінальною гілочкою аксона тієї нервової клітини, яка передає імпульс. Вона здебільшого розширена у вигляді гудзика або бутона квітки, вкрита пресинаптичною мембраною. У цьому полюсі розташовані мітохондріїта синаптичні пухирці, які містять так звані медіатори (нейротрансмітери). Останні сприяють передачі нервового імпульсу на постсинаптичну частину. Синаптичні пухирці різняться розмірами, ультраструктурою і хімічним складом: маленькі прозорі (30-60 нм), великі електронно-щільні (80-150 нм), прозорі пухирці з щільною гранулою (50-90 нм). Найпоширеніші нейромедіатори перераховані у табл. 22. Дофамін, гліцин та гамааміномасляна кислота є гальмівними медіаторами. Дослідженнями останніх років показано, що недостатня секреція дофаміну призводить до розвитку паркінсонізму. Пресинаптична мембрана містить електронно-щільні частинки діаметром 60 нм, які пов'язані між собою мікрофіламентами та утворюють пресинаптичну решітку для пухирців. Очевидно, остання визначає місця контакту синаптичних пухирців з пресинаптичною мембраною. Цитоплазматичний бік пресинаптичної мембрани містить також невеликі скупчення матеріалу середньої електронної щільності.
Постсинаптична частина синапса може містити значні скупчення електронно-щільного матеріалу. У такому разі вона відрізняється за зовнішнім виглядом від пресинаптичної частини (так звані асиметричні синапси). Електронно-щільний матеріал у постсинаптичній частині може також розміщатися окремими плямами, які нагадують топографію плям у пресинаптичній частині (симетричні синапси). Постсинаптична мембрана містить у своєму складі специфічний білок - рецептор медіатора, за участю якого реалізується вплив нейротрансмітера на постсинаптичну частину.
Синаптична щілина має розміри 20-30 нм, заповнена тканинною рідиною. Вона може містити електронно-щільні частинки або ниткоподібні структури, що розташовані на поверхні обох синаптичних мембран на зразок щетини у щітці. Можливо, така будова забезпечує утримання пре- і постсинаптичних мембран разом.
Таблиця 22. Найпоширеніші нейротрансмітери
|
Малі молекули |
Катехоламіни |
Нейроактивні пептиди |
|
Глютамат |
Дофамін |
Субстанція Р |
|
ГАМК (у-аміномасляна кислота) |
Норадреналін |
Енкефалін |
|
Гліцин |
Серотонін |
Ендорфін |
|
Ацетилхолін |
Гістамін |
Вазопресин |
|
Вазоактивний інтестинальний пептид |

Рис. 3.52. Синаптичні контакти: А - загальний принцип структурної організації синапса; Б - схема нейром'язового синапса з ілюстрацією механізму м'язового скорочення: нервовий імпульс зумовлює викидання ацетилхоліну з синаптичних пухирців у синаптичну щілину, що призводить до підвищення проникності мембран T-системи для іонів Са2+, які, у свою чергу, запускають механізм взаємного ковзання актинових та міозинових філаментів і скорочення саркомерів

Рис. 3.53. Трансмісійна електронна мікроскопія ділянки синаптичних контактів: А - синапс між симпатичним нервовим волокном та гладким міоцитом, х 45 000; Б - моторна бляшка (аксо-м'язовий синапс) у скелетному м'язі, х 33 000
Під час надходження нервового імпульсу до закінчення пресинаптичного нейрона синаптичні пухирці зливаються з пресинаптичною мембраною, їхній вміст виливається у синаптичну щілину і медіатор діє на постсинаптичну мембрану. Мембрана самих пухирців використовується повторно.
Функціонально розрізняють 2 види синапсів - збудливі та гальмівні. Морфологічні типи синапсів розрізняють залежно від того, які частини нейронів контактують між собою: аксодендритні (аксон першого нейрона передає імпульс на дендрит другого); аксосоматичні (аксон першого нейрона передає імпульс на тіло другого); аксо-аксонні (терміналі аксона першого нейрона закінчуються на аксоні другого). Вважають, що аксо-аксонні синапси виконують гальмівну функцію. Крім того, між деякими нейронами знайдені дендро-дендритні, а також дендросоматичні синапси. Таким чином, будь-яка частина нейрона може утворювати синапс з будь-якою частиною іншого нейрона.
Крім описаних хімічних, або відкритих, синапсів існують так звані електричні, або закриті, безпухирцеві синапси. Останні не мають синаптичної щілини, в них відсутні також синаптичні пухирці. У людини вони виявлені між нейронами мозочка.
Нейроглія. Нейрони існують у тісному генетичному, структурному та функціональному зв'язку з нейроглією. Цей термін, який був запропонований Р. Вірховим у 1846 р., означає у буквальному перекладі "нервовий клей", а насправді це середовище, що оточує нейрони. Побудована нейроглія з клітин. Її функції: опорна, розмежувальна, трофічна, секреторна, захисна.
Усі різновиди клітин нейроглії поділяють на дві групи: макроглію та мікроглію. У свою чергу, серед макрогліоцитів розрізняють епендимоцити, астроцити та олігодендроцити і, як різновид останніх у складі периферійної нервової системи, нейролемоцити (клітини Шванна) (рис. 3.49). Макроглія походить, як і нейрони, з нервової трубки, а мікроглія - із моноцитів і належить до макрофагічної системи (табл. 23).
Епендимоцити утворюють щільний, епітеліоподібний пласт клітин, які вистеляють спинномозковий канал і всі шлуночки мозку. Епендимоцити виникають першими у процесі гістогенезу нервової тканини з гліобластів нервової трубки. На цій стадії розвитку вони виконують розмежувальну й опорну функції. На поверхні клітин, звернених у порожнину каналу нервової трубки, містяться війки, яких може бути до 40 на одну клітину. Вважають, що війки сприяють рухові рідини у порожнинах мозку. Від базального кінця епендимоцита відходять довгі відростки, які розгалужуються і перетинають усю нервову трубку, утворюючи її опорний апарат. На зовнішній поверхні трубки ці відростки утворюють поверхневу гліальну пограничну мембрану, яка відмежовує нервову трубку від інших тканин.
Після народження епендимоцити виконують лише функцію вистилання порожнин мозку. Війки в епендимоцитах поступово втрачаються і зберігаються лише у деяких ділянках, наприклад, у водопроводі мозку. Деякі епендимоцити виконують секреторну функцію. Наприклад, епендимоцити субкомісурального органа продукують секрет, який, можливо, бере участь у регуляції водного обміну. Особливу будову мають епендимоцити, що вкривають судинні сплетення шлуночків мозку. Цитоплазма базального полюса цих клітин утворює численні глибокі складки, містить великі мітохондрії і різні включення. Епендимоцити судинних сплетень мозку беруть активну участь в утворенні спинномозкової рідини та регуляції її складу.
Таблиця 23. Походження та головні функції клітин нейроглії
|
Типи гліоцитів |
Походження |
Локалізація |
Головні функції |
|
Олігодендроцит |
Нервова трубка |
Утворення мієліну, електрична ізоляція |
|
|
Нейролемоцит (клітина Шванна) |
Нервова трубка |
Периферійна нервова система |
Утворення мієліну, електрична ізоляція |
|
Астроцит |
Нервова трубка |
Центральна нервова система |
Опорна, метаболічна, гематоенцефальний бар'єр, участь у відновних процесах |
|
Епендимоцит |
Нервова трубка |
Центральна нервова система |
Вистелення порожнин центральної нервової системи |
|
Мікроглія |
Стовбурові кровотворні клітини червоного кісткового мозку |
Центральна нервова система |
Макрофагічна активність |
Астроцити утворюють опорний апарат центральної нервової системи. Це невеликі клітини зірчастої форми з численними відростками, які розходяться у різні боки. Розрізняють протоплазматичні та волокнисті (фібрилярні) астроцити; існують також і перехідні форми астроцитів (волокнисто-протоплазматичні). Протоплазматичні астроцити локалізуються переважно у сірій речовині мозку. Розміри їх 15—25 мкм. Відростки короткі і товсті, розгалужені. На імпрегнованих солями металів препаратах ці клітини нагадують зарості чагарника. Волокнисті астроцити переважно розташовані у білій речовині мозку. Відростки їх довгі, прямі, слабко або зовсім нерозгалужені, на поперечному розрізі круглої або овальної форми.
Відростки астроцитів закінчуються на судинах, нейронах, базальній мембрані, яка відокремлює мозкову тканину від м'якої мозкової оболонки. У всіх випадках відростки розширюються на кінці і розплющуються на поверхні капіляра або нейрона, вкриваючи значну її частину й утворюючи так звану астроцитарну ніжку. Ніжки астроцитів контактують між собою і формують майже суцільну обгортку навколо капіляра або нейрона (лишаються вільними лише синаптичні контакти), входячи до складу гематоенцефального бар'єру.
Дослідженнями останніх років установлено, що клітини астроцитної глії гіпокампа та навколошлуночкової ділянки мозку стимулюють у дорослих ссавців утворення нейронів зі стовбурових нейральних клітин усіх мозкових ділянок.
У цитоплазмі астроцитів містяться фібрили, які складаються з мікрофіламентів. Кожний пучок мікрофіламентів починається у ніжці, йде через відросток до навколо-ядерного простору, а потім у другий відросток, доходячи до його кінця. Таким чином, цитоплазма астроцитів заповнена прямими або трохи звивистими пучками проміжних мікрофіламентів діаметром 8—9 нм. Очевидно, ці структури забезпечують міцність відростків астроцита. Ядро астроцита велике, світле. Цитоплазма також досить свила, тому що містить мало рибосом та елементів гранулярної ендоплазматичної сітки. У нервовій тканині трапляються астроцити, що дегенерують. Ймовірно, що процес загибелі і новоутворення астроцитів збалансований і популяція цих клітин може повільно відновлюватися
Олігодендроцити - найчисленніша група гліоцитів. Вони відрізняються невеликими розмірами, наявністю коротких, дуже тонких відростків. Тіла їх мають багатокутну або овальну форму. Олігодендроцити оточують тіла нейронів та їхні відростки на всій довжині, локалізуються як у центральній, так і периферійній нервовій системі. Щільність цитоплазми клітин олігодендроглії наближається до цього показника нервових клітин. Цитоплазма олігодендроцитів не містить нейрофіламентів. Функції цих клітин дуже різноманітні: трофічна, ізолювальна, участь у водно-сольовому обміні, процесах дегенерації і регенерації нервових волокон. Олігодендроцити, які утворюють оболонки навколо відростків нервових клітин, мають назву нейролемоцитів (клітин Шванна).
Мікроглія - сукупність маленьких клітин з двома-трьома відростками, які мають на своїй поверхні короткі вторинні і третинні розгалуження. Ядра клітин мікроглії витягнутої або трикутної форми, багаті на гетерохроматин. У разі подразнення нервової тканини (запалення, рана) клітини мікроглії змінюються - збільшується об'єм ядра і цитоплазми, клітини стають круглими, рухомими, втягують свої відростки. Подібно до інших макрофагів мікрогліоцити наповнюються фагоцитованим матеріалом. У такому вигляді їх називають зернистими кулями. Останнім часом показана здатність мікроглії брати участь у синтезі білків-імуноглобулінів (антитіл).
Нервові волокна - це відростки нервових клітин, укриті оболонками. Залежно від будови оболонки вони поділяються на дві основні групи - мієлінові та безмієпінові (рис. 3.54, 3.55, 3.56, 3.57). І ті, й інші побудовані з осьового циліндра, який є відростком нервової клітини та оболонки, утвореної клітинами олігодендроглії (нейролемоцитами).
Мієлінові нервові волокна мають досить складну будову. Вони є як у центральній, так і в периферійній нервовій системі, тобто у складі головного і спинного мозку, а також у складі периферійних нервів. Це товсті волокна, діаметр їхнього поперечного перерізу коливається від 1 до 20 мкм. Вони побудовані з осьового циліндра, мієлінової оболонки, нейролеми та базальної мембрани. Осьовий циліндр - це відросток нервової клітини, яким частіше буває аксон, але може бути і дендрит. Він складається в нейроплавми, яка містить поздовжньо орієнтовані нейрофіламенти і нейротубули, а також мітохондрії. Осьовий циліндр укритий аксолемою (продовженням клітинної мембрани), яка забезпечує проведення нервового імпульсу.
Мієлінова оболонка - це муфта товщиною від 0,3 до 15-20 мкм, яка покриває осьовий циліндр уздовж. Вона відсутня у місці відходження відростка від перикаріона, в ділянці термінальних розгалужень аксона і в ділянках, які мають назву вузлів (перетяжок) Ранв'є (рис. 3.54). Ділянка волокна між двома сусідніми вузлами називається міжвузловим сегментом. Довжина останнього - від кількох мікрометрів до кількох міліметрів. Вузлова перетяжка має розміри 0,25-1 мкм.
Мієлінова оболонка містить Ліпіди і тому фарбується у чорний колір у разі обробки осмієвою кислотою. На певній відстані одна від одної у темній мієліновій оболонці розташовуються вузькі світлі лінії, що йдуть у косому напрямку. Це так звані насічки мієліну Шмідта-Лантермана. За допомогою електронного мікроскопа було виявлено, що мієлінова оболонка має пластинчасту будову. Остаточно зрозуміти будову мієлінової оболонки допомогли дослідження процесу розвитку мієлінових нервових волокон.
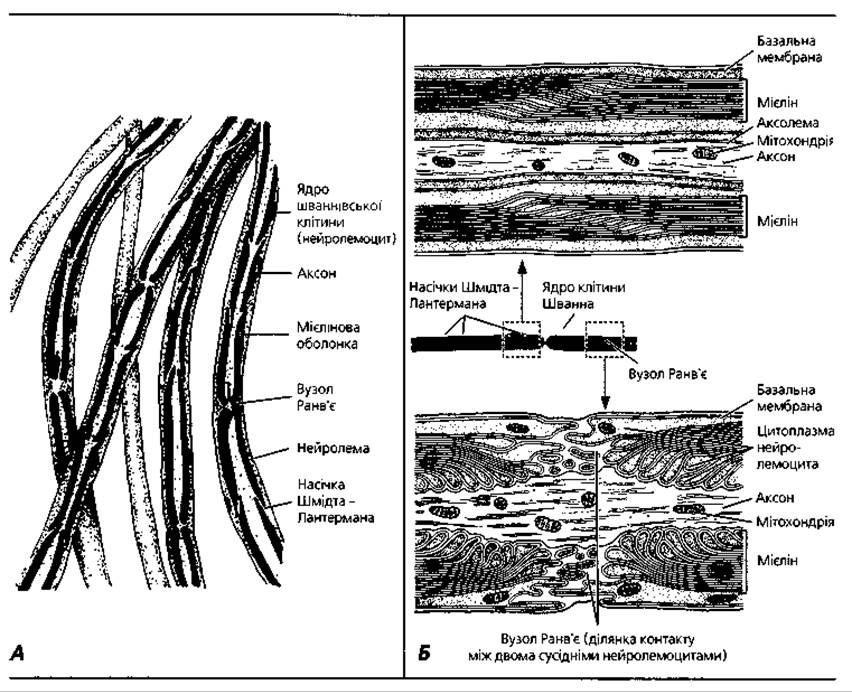
Рис. 3.54. Мієлінові нервові волокна: А - напівсхематичне відтворення розщипаного гістологічного препарату, імпрегнація осмієм, х 600; Б - схема, що ілюструє будову вузлів Ранв'є та насічок Шмідта-Лантермана

Рис. 3.55. Трйвимірна реконструкція нервових волокон: А - послідовні фази формування мієлінової оболонки; Б - ділянка вузла Ранв'є мієлінового нервового волокна; В - безмієлінове нервове волокно з ізольованими аксонами; Г - безмієлінове волокно кабельного типу

Рис. 3.56. Схеми ультраструктурної організації різних типів нервових волокон: А - мієлінові волокна центральної нервової системи: один олігодендроцит формує мієлінові оболонки навколо 3-50 нервових волокон. Мієлінове (Б) та безмієлінове (В) волокна периферійної нервової системи: оболонку навколо аксонів формують нейролемоцити (клітини Шванна)

Рис. 3.57. Електронна мікроскопія нервових волокон: А - мієлінове нервове волокно, поперечний зріз, х 35 000; Б - мієлінові та Безмієлінові нервові волокна у складі фрагмента периферійного нерва, х 20 000
У процесі розвитку мієлінового волокна осьовий циліндр занурюється у нейролемоцит, угинаючи його оболонку та утворюючи глибоку складку (рис. 3.55, А). Ця подвійна складка (дуплікатура) плазмолеми нейролемоцита отримала назву мезаксона. У процесі подальшого розвитку нейролемоцит повільно обертається навколо осьового циліндра, внаслідок чого мезаксон багато разів огортає його. Цитоплазма нейролемоцита і його ядро лишаються на периферії, утворюючи нейролему. Таким чином, мієлінова оболонка утворюється із щільно концентрично нашарованих навколо осьового циліндра завитків мезаксона, які і є пластинками мієлінового шару.
Кожний завиток мезаксона має ширину близько 8-12 нм і відповідає ліпідним шарам двох листків плазмолеми нейролемоцита. На його середині та на поверхні під електронним мікроскопом можна спостерігати тонкі темні лінії, утворені білковими молекулами. Насічки мієліну відповідають тим місцям, де завитки мезаксона розсунуті цитоплазмою нейролемоцита. Оболонку одного нервового волокна утворюють багато нейролемоцитів. Вони контактують між собою у ділянках вузлових перетяжок. Міжвузловий сегмент відповідає території, укритій одним нейролемоцитом.
На поздовжньому розрізі мієлінового нервового волокна поблизу вузла Ранв'є є ділянка, в якій завитки мезаксона послідовно контактують з осьовим циліндром. Місця прикріплення найглибших завитків найбільш віддалені від вузла, а всі наступні - поступово наближуються до нього. Це пояснюється тим, що мезаксон нашаровується у процесі росту і осьового циліндра, і нейролемоцитів. Таким чином, перші шари мезаксона коротші, ніж останні. Краї двох суміжних нейролемоцитів, контактуючи у ділянці перетяжки, утворюють пальцеподібні вирости діаметром біля 50 нм. Довжина виростів різна. Разом вони мають характерний вигляд "пишного комірця".
Нейролема - тонка, світла у разі обробки осмієвою кислотою оболонка нервового волокна, розташована ззовні від шару мієліну. Нейролема утворена цитоплазматичними частинами нейролемоцитів і їхніми ядрами. Базальна мембрана, вкриваючи ззовні нервове волокно, сполучається з колагеновими волокнами ендоневрію (сполучною тканиною, що оточує нервові волокна).
Вищеописану будову мають мієлінові нервові волокна периферійної нервової системи. Мієлінові волокна центральної нервової системи мають низку особливостей: їхню оболонку замість нейролемоцитів утворюють типові олігодендроцити (в останніх менше цитоплазми, вони дрібніші); відсутні насічки мієліну і базальна мембрана; вузлові перетяжки мають більші розміри, а міжвузлові сегменти коротші, окрім того, один олігодендроцит може одночасно формувати оболонку навколо кількох нервових відростків (рис. 3.56, А).
Безмієлінові нервові волокна є типовими для автономного відділу нервової системи. Діаметр волокон 1-4 мкм, тобто вони тонші, ніж мієлінові волокна. Складаються безмієпінові волокна з осьового циліндра, нейролеми і базальної мембрани. Нейролема утворена тяжем нейролемоцитів, які щільно прилягають один до одного. Прогинаючи оболонку нейролемоцитів, осьовий циліндр глибоко занурюється у цей тяж, а гліальна клітина як муфта огортає відросток. Оболонка нейролемоцита утворює глибоку складку, мезаксон, на зразок того, що вже описаний вище для мієлінового волокна. Якщо тяж лемоцитів охоплює не один осьовий циліндр, а кілька (10-20), то такі безмієпінові волокна називають поліаксонними, або волокнами кабельного типу. Ззовні безмієпінове нервове волокно, як і мієлінове, вкрите базальною мембраною (рис. 3.55, 3.56, 3.57).
Швидкість передачі нервового імпульсу мієліновими нервовими волокнами значно вища (5-120 м/с), ніж безмієліновими (1-2 м/с). Це пояснюється тим, що у безмієпіновому волокні хвиля деполяризації рухається по всій плазмолемі не перериваючись, а у мієліновому вона йде сальтаторно, тобто стрибками, виникаючи лише у ділянках вузлів Ранв'є.
Нервова тканина розвивається з нервової пластинки, яка є потовщенням ектодерми на спинному боці зародка. Нервова пластинка послідовно перетворюється у нервовий жолобок і нервову трубку, яка, замикаючись, відокремлюється від шкірної ектодерми (див. розділ 2). Частина клітин нервової пластинки лишається між нервовою трубкою і шкірною ектодермою у вигляді пухкого скупчення клітин, так званого нервового гребеня, або гангліозної пластинки. Клітини гребеня мігрують у латеральному і вентральному напрямках і дають такі похідні: ядра черепних нервів, нейрони спинномозкових і автономних вузлів, нейролемоцити (нейроглію), пігментні клітини шкіри.
Клітини нервової трубки, яка побудована з багаторядного нейроепітелію, мають назву вентрикулярних, або нейроепітеліальних, клітин. Вони циліндричної форми, (хні апікальні частини межують з порожниною нервової трубки і сполучені щілинними контактами, а базальні - контактують з субпіаль- ною пограничною мембраною, їм властивий процес циклічного переміщення ядер. Здатність цих клітин до розмноження зменшується у процесі ембріонального розвитку і після народження. Морфологічно подібні вентрикулярні клітини шляхом диференціації перетворюються у різні типи клітин нервової тканини. Частина з них дає початок нейронам, інша - гліальним клітинам (епендимоцитам, астроцитам, олігодендроцитам). У деяких ділянках мозку з вентрикулярних клітин утворюються так звані субвентрикулярні й екстравентрикулярні нейрогермінативні клітини, які довше зберігають проліферативну активність й існують ще деякий час після народження. З них утворюються деякі типи нейронів і нейроглії.
Терміни для запам'ятовування
1. Нервова тканина. 2. Нейрон (нейроцит). 3. Перикаріон. 4. Аксон. 5. Дендрит. 6. Хроматофільна субстанція (речовина Ніссля, тигроїд). 7. Нейрофібрила. 8. Аксонний транспорт. 9. Дендритний транспорт. 10. Уніполярний нейрон. 11. Біполярний нейрон. 12. Псевдоуніполярний нейрон. 13. Мультиполярний нейрон. 14. Аферентний (рецепторний) нейрон. 15. Асоціативний (вставний) нейрон. 16. Еферентний (моторний) нейрон. 17. Рефлекторна дуга. 18. Синапс. 19. Пресинаптична частина. 20. Постсинаптична частина. 21. Синаптична щілина. 22. Аксосоматичний синапс. 23. Аксодендритний синапс. 24. Аксо-аксонний синапс. 25. Дендро-дендритний синапс. 26. Дендросоматичний синапс. 27. Нейроглія. 28. Макроглія. 29. Епендимоцит. 30. Астроцит. 31. Протоплазматичний астроцит. 32. Волокнистий астроцит. 33. Олігодендроцит. 34. Нейролемоцит (клітина Шванна). 35. Мікроглія. 36. Нервове волокно. 37. Мієлінове нервове волокно. 38. Мієлінова оболонка. 39. Вузол (перетяжка) Ранв'є. 40. Міжвузловий сегмент. 41. Насічка мієліну. 42. Мезаксон. 43. Нейролема. 44. Безмієлінове нервове волокно. 45. Нервова пластинка. 46. Нервовий жолобок. 47. Нервова трубка. 48. Нервовий гребінь (гангліозна пластинка). 49. Вентрикулярна (нейроепітеліальна) клітина. 50. Екстравентрикулярна (нейрогермінативна) клітина.
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.