Фізіологія рослин - Мусієнко М.М. 2001
Фізіологія росту та розвитку рослин
Проростання. Первинний та вторинний ріст
Проростання
Ріст рослин розпочинається з проростання насіння. Період проростання є важливим етапом онтогенезу рослини, що має великий вплив на його подальший ріст, розвиток та продуктивність. Встановлено, що кінцевий врожай краще корелює з ранньою появою сходів в порівнянні з іншими параметрами. Практикою рослинництва показано, що якісне насіння швидко і дружно проростає. Але у більшості випадків насіння різних видів сільськогосподарських культур проростає нерівномірно, що зумовлено його різною якістю, яка встановлюється при дозріванні. Чим довший період від посіву до появи сходів, тим більша ймовірність пошкодження проростаючої насінини мікроорганізмами та грибами, що живуть у грунті. Тому широко досліджуються способи застосування фізіологічно активних речовин, мікроелементів, вітамінів та інших сполук, а також фізичних факторів з метою підвищення швидкості проростання насіння. Ця ознака важлива і для селекційної роботи.
На даний час встановлено, що надійним критерієм проростання є не прокльовування насінини, як це вважалося раніше, а ВСТУП клітин апікальних меристем у фазу синтезу ДНК та мітоз, оскільки при певних умовах корінець може проклюнутись, а проросток не формується. Звідси витікає, що часові параметри першого клітинного циклу в меристемі кореня, зокрема, таких його маркерних процесів, як Синтез ДНК та мітоз, можуть мати практичне значення для визначення фізіологічної активності насіння.
Процес проростання включає в себе і ті процеси, які проходять в насінині до того, як проявляються ознаки видимого росту. У покритонасінних насінина складається із зародка, ендосперму та насінної шкірки. Залежно від тих чи інших запасних речовин розрізняють насіння, яке запасає олії, крохмаль або білки. Насіння всіх рослин має фітин. Головна його функція — забезпечення зародка сполуками фосфору. Одночасно фітин містить деяку кількість К+, Мg2+ та Са2+. В насінні є також Ферменти і гормони, однак в неактивному стані. Розподіл речовин в насінні нерівномірний. Так, Тканини зародка збагачені мінеральними елементами, амінокислотами, ферментами, гормонами, причому особливо значний вміст їх в зародковому корінчику.
Деякі ферменти знаходяться в зв’язаному, неактивному, стані і лише під впливом набрякання переходять в активний, стан. Однак, ряд ферментів синтезується de novo. Для цього потрібні інформаційні РНК.
За часом утворення інформаційну РНК в проростаючому насінні поділяють на три типи. До першого типу відносять, так звану, попередню, або залишкову іРНК, яка була транскрибована з ДНК ще в період ембріогенезу насіння. Однак, її вміст незначний, тому і роль невелика. До другого типу відносять РНК, яка також транскрибована в ембріогенезі, однак яка не пройшла процесингу і тому неактивна. Під час набрякання в ній відбуваються необхідні перетворення і забезпечується Синтез білків, специфічних для проростання, зокрема ферментів гідролаз. Нарешті, третій тип— це новоутворена РНК, яка з’являється через 1-2 години після намочування. Ця РНК транскрибується під час проростання з ДНК, за участю РНК-полімерази І, і відповідає за синтез специфічних білків — ферментів. Вважають, що в синтезі білка при проростанні спочатку беруть участь Рибосоми, утворені ще в ембріогенезі, потім, десь через 8 годин після намочування насіння спостерігається посилене утворення рибосомальної РНК і формуються нові рибосоми.
Велику роль в регуляції утворення ферментів відіграють фітогормони, зокрема Гібереліни. Джерелом гіберелінів е зародок. В сухому насінні гібереліни знаходяться в зв’язаному стані. Вони активуються та частково утворюються заново під впливом набрякання. Так як гідролази, зокрема нуклеази, каталізують розпад нуклеїнових кислот, з’являються пуринові основи, які дають початок фітогормону цитокініну. Одночасно під впливом протеаз білки розпадаються до амінокислот, серед яких є Триптофан, який є попередником фітогормону ауксину. Цитокініни та Ауксини регулюють ріст зародка.
Росту органів зародка сприяють і ферменти пектиназа і целюлаза. Ендорамнази розщеплюють поліцукри (маннани), які з’єднують фібрили целюлози, чим дають змогу зародковій осі подолати опір ендосперму. Абсцизова кислота гальмує утворення ферменту маннази, що і є однією з причин гальмівної дії даного ферменту на проростання насіння. Для того щоб розпочався поділ ядра, необхідна редуплікація ДНК. Синтез ДНК в проростаючому насінні розпочинається значно пізніше за Синтез РНК та білків. Характерно, що в сухому насінні основна частина клітин знаходиться в стані (передсинтетичний період).
Для розвитку зародка, що знаходиться в насінині, потрібні конкретні умови середовища. Перш за все потрібна Вода. Повітряно сухе насіння містить від 5 до 15% води і перебуває у стані вимушеного спокою. Однак досить часто, навіть при наявності певних оптимальних умов, насіння не проростає, доки в ньому не відбудуться певні зміни, пов’язані з фізіологією дозрівання. Іншими словами, в насінині завжди є механізми, які забезпечують збігання термінів проростання з надходженням сезону, який сприяв би росту даного організму.
В цих механізмах вагому роль відіграють зовнішні оболонки насінин, які часто не проникні для води, кисню, а інколи містять і інгібітори росту. В такому насінні механічне пошкодження оболонок (скарифікація) може прискорити процеси проростання В природних умовах аналогічні ефекти можуть виконувати різноманітні бактерії, які руйнують оболонку насінини. Деякі плоди та насіння містять інгібітори росту. Найчастіше — це абсцизова кислота. Можливо, при довготривалому намочуванні насіння інгібітори вимиваються і гальмуючий вплив їх зникає. Паралельно цьому процесу може зростати вміст стимуляторів росту, наприклад, гіберелінів.
Нарешті, в процесах проростання важливу роль відіграють Температура. Оптимальні температури для проростання насіння звичайно відповідають тим, які характерні для ареалу поширення даного виду рослин. Для того щоб відбулося проростання насіння деяких видів рослин, воно повинно перенести вплив понижених температур.
Є рослини, для проростання насіння яких потрібне світло. Після набрякання насіння світло, у деяких видів рослин, часто перериває спокій насіння і цей процес контролюється фітохромами.
Після періоду спокою розпочинається розвиток зародка. Для цього відбувається адсорбція води крізь мікропіле та насінні оболонки за рахунок колоїдів, якими є білки, крохмаль, геміцелюлози, пектинові речовини. Набрякання цих речовин створює таку силу, яка достатня для розривання оболонки, що оточує насінину. Далі шляхом осмосу вода пересувається від клітини до клітини, і в процесі транспортування вона активує цілий ряд біохімічних процесів. Насамперед вода бере участь у гідролізі запасних поживних речовин.
Для нормального проростання потрібні певні температури, які впливають на Швидкість ферментативних реакцій, а також вільний доступ кисню. Кисень потрібний для забезпечення процесів аеробного Дихання. Фізіологія проростання пов’язана з функціонуванням двох зон активності: це — запасні речовини та область зародка. Процеси розкладання поживних речовин нам уже відомі, тому, не звертаючи уваги на процеси гідролізу, скажемо, що всі продукти розкладання транспортуються до зони росту зародка. Енергія, потрібна для синтетичних процесів, надходить за рахунок дихання. Все це призводить до того, що різко зменшується суха маса насінини, і цей процес продовжується доти, доки в проростку не з’являться зелені листки, і не розпочнеться автотрофне живлення.
Ріст зародка відбувається шляхом поділу клітин та диференціювання їх. Перша ознака росту — це поява зародкового корінчика. Він характеризується позитивним геотропізмом, тому росте вниз, цим самим закріплюючи насінину в грунті. Потім з’являється пагін-плюмула, або брунечка, яка має від’ємний геотропізм, тобто росте вгору. Розрізняють два типи проростання: залежно від того, чи сім’ядолі залишаються під землею, чи виносяться на поверхню грунту (рис. 159).

Рис. 159. Етіольовані підземні проростки й зміни їхньої форми, індуковані світлом при переході до надземного способу життя: а — злаки; б — дводольні з гіпогеальним проростанням; в — дводольні з епіґеальним проростанням: 1 — колеоптиль, 2 — мезокотиль. З — первинний складений листок, 4 — епікотиль, 5 — гіпокотиль
У дводольних при витягуванні міжвузля, яке знаходиться безпосередньо під сім’ядолями (гіпокотиль), сім’ядолі виносяться на поверхню. Таке проростання називають епігеальним. Якщо ж міжвузля, що знаходиться над сім’ядолями (епікотиль), витягується, то сім’ядолі залишаються під землею і такий тип проростання називають гіпогеальним. В обох випадках зігнута частина міжвузля захищає сім’ядолю від руйнування, а, з’явившись на світло, ця ділянка відразу ж випрямляється. Цей процес контролюється фітохромом. У злакових рослин, які теж належать до однодольних, плюмула захищена чохликом — колеоптилем, який характеризується позитивним фототропізмом та від’ємним геотропізмом.
Перший листок з’являється через колеоптиль і швидко розпускається під впливом світла (рис. 160). Світло спричинює ряд реакцій, що контролюються фітохромом і мають назву фото- морфогенез. Ці зміни зумовлюють перехід етіольованого росту до нормального. Сім’ядолі перетворюються на справжні листки. Відбувається синтез хлорофілів. Проросток переходить до автотрофного типу живлення.
Таким чином, для нормального росту та розвитку проростків потрібне світло. Як тільки з’являються перші зелені листочки, розпочинається Фотосинтез. На цьому фаза проростка закінчується, а рослина переходить в ювенільну фазу.
На світлі у мезофілі листка із пропластид розвиваються Пластиди. Причому, ще до завершення росту пластид та формування гранулярно-ламелярної структури розпочинається синтез ферментів, які каталізують темнову стадію фотосинтезу. Фермент карбоксилування РУБІСКО присутній в проростках ще до того, як листя набуває зеленого забарвлення. В той же час на світлі відбувається новоутворення цього ж ферменту. Для початку фотосинтезу необхідна також наявність первинного акцептора СО2 рибулозо-1,5-бісфосфату. Він утворюється як проміжний продукт пентозофосфатного шляху дихання. Відомо, що ще до проростання в зародку присутній головний фермент цього шляху дихання — глюкозофосфатдегідрогеназа. Під час проростання активність даного ферменту різко зростає, що і спричинює нагромадження акцептора СО2. Коли розпочинає роботу цикл Кальвіна, активність глюкозофосфатдегідрогенази падає.
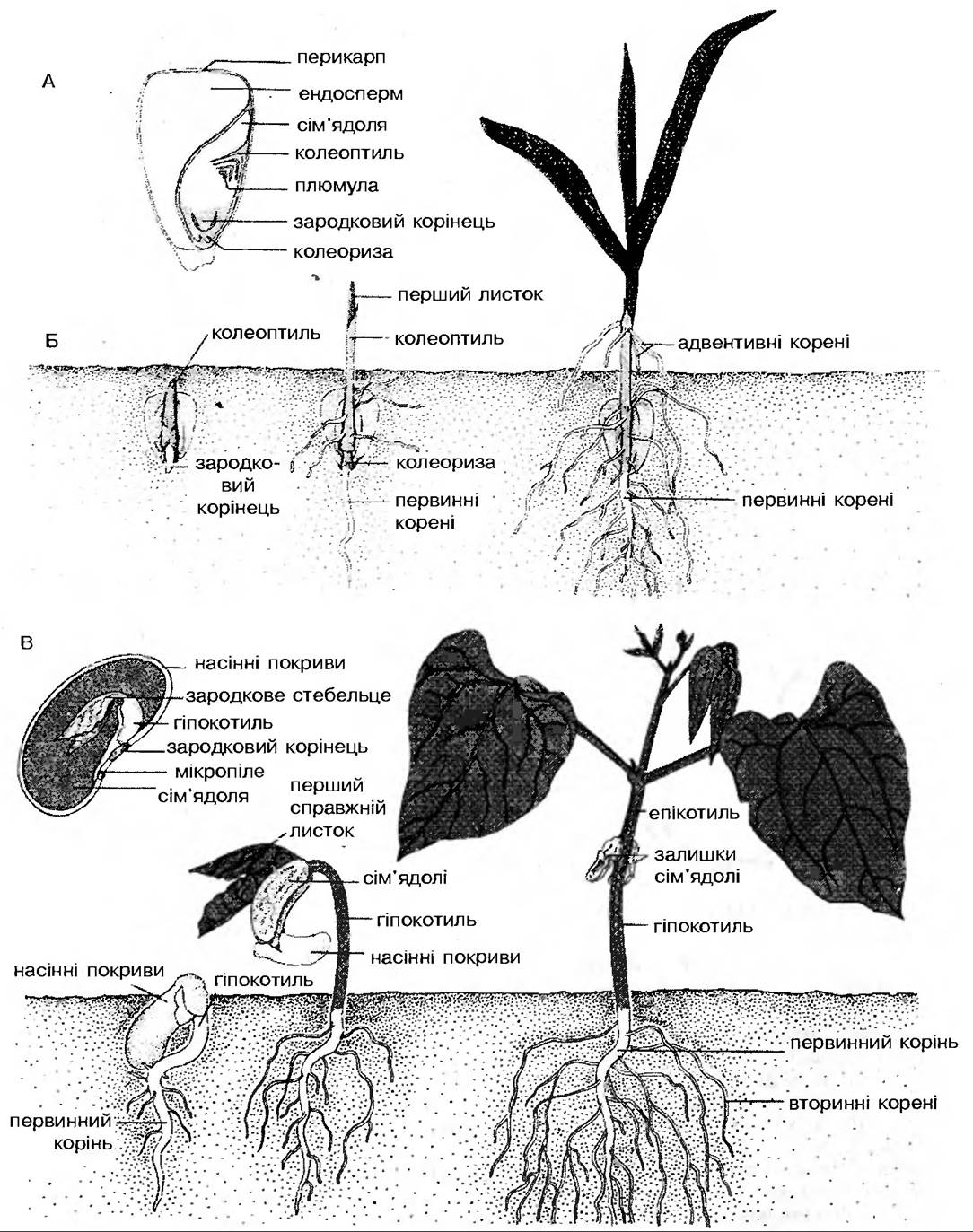
Рис. 160. Проростання насінини (А) і розвиток проростку (Б) однодольних та дводольних (В) рослин
Таким чином, проростання розпочинається з набрякання. У деяких видів уже при цьому лопається насіннєва шкірка. Наступний етап гідролізу та транспорту запасних речовин супроводжується інтенсивним диханням. Потім завдяки росту клітин розтягуванням і відновленню поглинання води збільшується зародок. Нарешті, з виходом зародкового корінця при розриві насіннєвої шкірки, насінина вважається пророслою.
Проростання насіння залежить від багатьох умов. В природі рідко всі вони бувають оптимальними, тому проростання насіння навіть одного й того ж виду може розтягнутися на роки. Це дає змогу зберегти життєдіяльність певної кількості насіння навіть тоді, коли несприятливі умови зумовлять загибель пророслого насіння. В цілому ж, проростання насіння є одним із критичних етапів в житті рослинного організму, від якого значною мірою залежить виживання певного виду.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.