Фізіологія рослин - Мусієнко М.М. 2001
Загальні принципи регуляції росту та морфогенезу рослин
Цитокініни
Відкриття цитокінінів повязане з дослідженнями культури тканин. Ще в 1950-1955 рр. було виявлено, що клітини калусу в стерильній культурі через певний проміжок росту на поживному середовищі припиняють Поділ клітин.
С. Міллер та Ф. Скуг (Вісконський університет, США) намагалися зумовити поділ зрілих серцевинних клітин тютюну in vitro. В інтактних рослинах ці клітини збільшуються в розмірах, але ніколи не діляться. Однак, якщо в середовище ввести кокосове молоко, вони починають
ділитися. Випробовуючи різні сполуки, Ф. Скуг помітив, що аденін при наявності ауксинів дещо стимулює клітинний поділ. Потім було перевірено НУКЛЕЇНОВІ КИСЛОТИ, до складу яких входить аденін, і виявлено, що додаючи до поживного середовища похідні ДНК, можна відновити поділ клітин.
В 1956 р. було виділено активне начало (6-фурфуриламінопурин), що викликає поділ клітин, яке одержало назву кінетин. Хоча кінетин не виявлено в рослинних тканинах, виявилось, що речовини з подібною біологічною активністю широко поширені у рослин. Їх назвали цитокініни. Цей термін має фізіологічний, а не хімічний зміст. Цитокінін являє собою речовину, що стимулює поділ клітин в культурі калусної Тканини рослин, яка росте на поживному середовищі з необхідними речовинами органічного і мінерального походження та екзогенний ауксин в оптимальній концентрації. Лише в 1963 р. вперше було ідентифіковано цитокінін, який виділили з недозрілого насіння кукурудзи і назвали зеатин. Пізніше Захау (1966), а потім і Скуг встановили, що цитокініни присутні у всіх видів тРНК. Цитокініни виявлено в самих різноманітних рослинних тканинах та особливо багаті на них верхівки коренів, ксилемний сік, проростаюче насіння, дозріваючі плоди, пухлини корончатих галів, що виникають при зараженні Agrobacterium tumefaciens. Досить високий вміст цитокінінів в кореневих бульбочках азотфіксуючих організмів.
Подібні речовини виявлені в різних мікроорганізмів, водоростей, папоротей, мохів, хвощів та вищих рослин. Зеатин вважають найактивнішим із природних цитокінінів, хоча тепер вже синтезовано і більш активні сполуки (рис. 177).
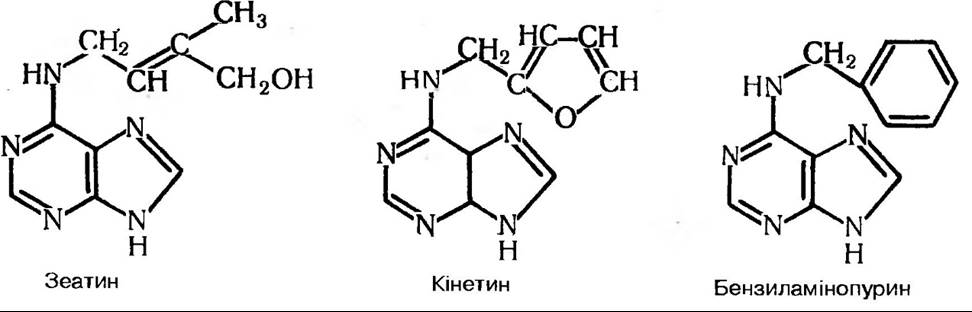
Рис. 177. Цитокініни
Шлях біосинтезу зеатину в рослинному організмі такий:
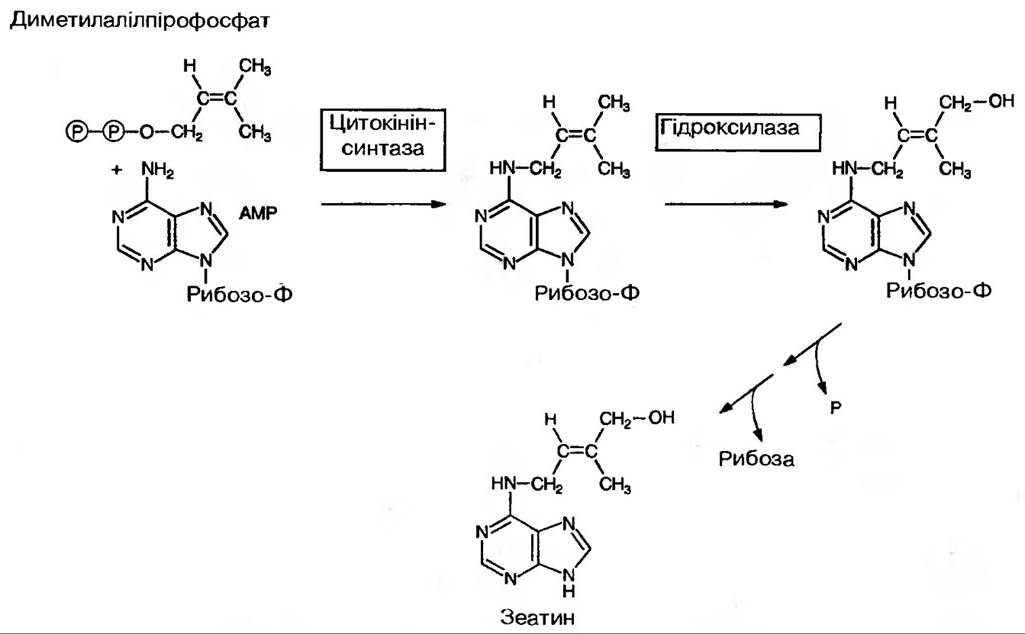
Найбільш багаті на них недозрілі плоди та насіння, а також меристеми. Основне місце синтезу — апікальні меристеми кореня. Є дані, що вони синтезуються також в дозріваючих плодах та насінні. Цитокініни, які присутні в тРНК, синтезуються у всіх живих клітинах рослин, в таких їхніх компартментах як Цитоплазма і хлоропласти, а можливо і в мітохондріях. Із кореня цитокініни, найбільш ймовірно пасивно транспортуються в надземні органи по ксилемі. Щодо цитокінінів, які синтезуються в інших частинах рослини, то вони, можливо, і не транспортуються.
Подібно аденіну, природні цитокініни зустрічаються в формі рибонуклеозидів та рибонуклеотидів , які, можливо, є попередниками вільних цитокінінів. Зв’язані цитокініни у вигляді риботидів чи рибозидів являють собою транспортні та запасні форми. В наукових дослідженнях широко використовуються синтетичні цитокініни типу кінетину та бензиладеніну, бо чим активніший природний цитокінін, тим важче його одержати.
Шляхи синтезу цитокінінів фактично мало вивчені. Рослинні тканини здатні розщеплювати цитокініни, що викликає втрату їх активності. Фермент, який каталізує даний процес, виділений із насіння кукурудзи; він активний лише в присутності кисню, тому йому присвоєна загальна назва цитокінін-оксидаза.
існують різноманітні біотести цитокінінів, які основані на:
·здатності цитокінінів стимулювати ріст розтягуванням у ізольованих листків або сім’ядолей, наприклад у редиски;
·стимуляції росту водяних рослин, наприклад Lemnaminor;
·прискоренні росту відрізків стебла або колеоптилей;
·здатності цитокінінів індукувати поділ клітин в культурі клітин;
·на специфічній властивості цитокінінів затримувати процеси Старіння листків;
·здатності цитокінінів стимулювати біосинтез пігментів, наприклад хлорофілу в сім’ядолях огірків.
Фізіологічна дія цитокінінів
Відомо різноманітні фізіологічні ефекти цитокінінів, проте всі їх можна умовно розділити на дві групи:
·стимуляція поділу клітин та їх диференціювання,
·затримка процесів старіння.
Цитокініни індукують поділ клітин, однак обов’язково при наявності ауксинів (рис. 178).
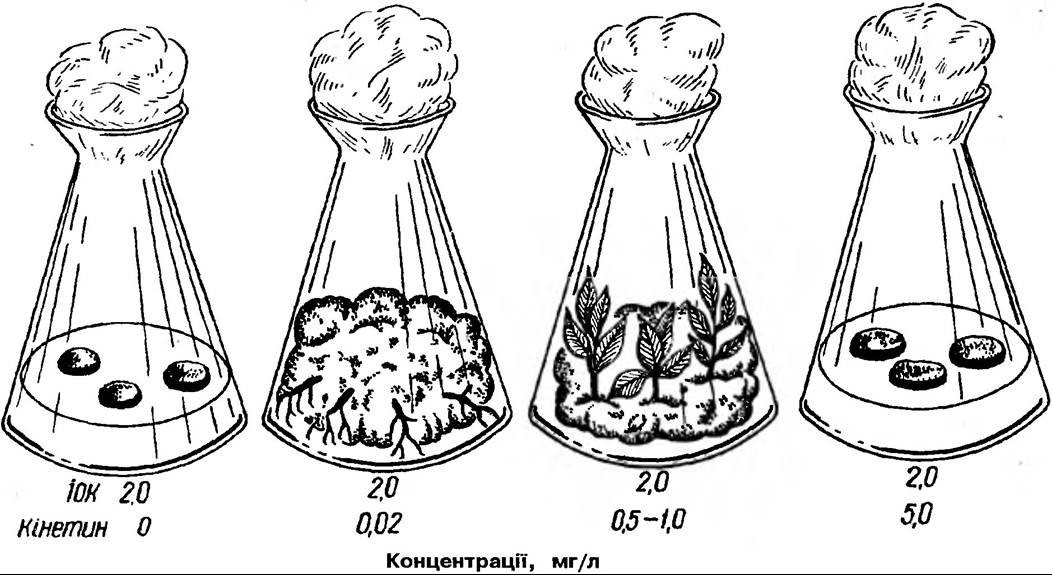
Рис 178. Спільна дія 10К, (2 мг/л) та кінетину на ріст і процеси диференціювання калуса серцевини стебла тютюну
Вони діють як специфічні стимулятори мітозу. Цитокініни відіграють істотну роль в процесах Диференціювання клітин та органогенезу в культурі тканин. Так, в калусній тканині серцевини тютюну відповідні концентрації ауксинів та кінетину зумовлюють утворення коренів або бруньок. При високих концентраціях ауксину формуються корені, а кінетину — бруньки. Якщо обидва гормони в однаковій кількості, в калусі формуються недиференційовані клітини.
В основі ростових ефектів цитокінінів лежить насамперед процес прискорення клітинного поділу, пов’язаний з посиленим синтезом ДНК. Вони індукують утворення бруньок, можуть спричинити розпускання бруньок, загальмованих ауксином, а також проростання насіння.
Під впливом цитокінінів відбувається загальна стимуляція обміну речовин, насамперед, синтезу РНК та білків. Цитокініни затримують старіння, тому що вони сприяють процесу підтримки певного рівня білків та нуклеїнових кислот, можливо, шляхом зниження швидкості розкладання їх, а також в результаті збереження цілісності клітинної мембрани.
Подібно ауксинам, цитокініни проявляють атрагуючу дію, тобто посилюють пересування речовин до збагачених ними тканин.
Цитокініни відвертають розпад хлорофілу та деградацію структур в ізольованому листку. Вони впливають на ультраструктуру хлоропластів, утворення в них мембран і гран, тому їх присутність необхідна умова нормального розвитку листка та підтримки його атрагуючої здатності Оброблені кінетином листки залишаються зеленими впродовж тривалого часу. Інтенсивність фотосинтезу у оброблених цитокінінами листках зростає. Активується синтез ключового ферменту темнових реакцій фотосинтезу — рибулозобісфосфаткарбоксилази, посилюється інтенсивність фотофосфорилування.
На молекулярному рівні цитокініни в комплексі з специфічним білковим рецептором посилюють активність РНК-полімерази та матричну активність хроматину. Це зумовлює збільшення кількості полірибосом та активацію синтезу білків, в тому числі деяких ферментів (наприклад, нітра- тредуктази). Відомо, що кінцева дія цитокінінів спричинює зміну експресії гена, можливо, на транскрипційному рівні. Є також дані про їхній вплив на транспортування іонів калію, Н+ та Са2.
Оскільки застосування цитокінінів займає важливе місце в експериментах з культурою тканини, вони мають важливе значення для різних біотехнологічних процесів.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.