Цитологія, загальна гістологія та ембріологія - В. К. Напханюк 2002
Тканини
Тканини внутрішнього середовища
Гемоцитопоез
Гемоцитопоез (haemocytopoesis) — це процес постійного утворення та розвитку формених елементів крові, внаслідок чого підтримується постійність їх кількісного та якісного складу.
Оскільки в результаті кровотворення компенсується природна втрата віджилих формених елементів, гемоцитопоез можна вважати процесом фізіологічної регенерації крові.
Розрізняють ембріональний гемоцитопоез, який відбувається в ембріональному періоді, і постембріональний, який є процесом фізіологічної регенерації крові.
Ембріональний гемоцитопоез
Це процес утворення крові як Тканини.
Починається ембріональне кровотворення на другому тижні внутрішньоутробного розвитку в стінці жовткового мішка, де з мезенхіми утворюються Кров’яні острівці, в центрі яких виникають стовбурові клітини крові. Периферійні клітини кров’яних острівців випинаються, з’єднуються між собою, утворюючи ендотеліальну вистілку кров’яних судин. Кровотворення відбувається інтраваскулярно, при цьому утворюються первинні еритроцити — мегалоцити. Цей тип кровотворення має назву мегалобластичного кровотворення, він є характерним тільки для ембріонального періоду розвитку, але може виявлятися і в постнатальному періоді при деяких захворюваннях (злоякісні анемії). Паралельно з мегалобластичним у стінці жовткового мішка починається нормобластичне кровотворення, при якому з бластів утворюються вторинні еритробласти, які перетворюються на поліхроматофільні еритробласти, далі на нормобласти, з яких утворюються вторинні еритроцити (нормоцити). Водночас із бластів, розташованих екстраваскулярно (навколо судинних стінок) диференціюється невелика кількість гранулоцитів — нейтрофілів і еозинофілів. Частина стовбурових клітин залишається в недиференційованому стані і розноситься з кров’ю до різних органів зародка. Після редукції жовткового мішка основним кровотворним органом стає Печінка.
Кровотворення у печінці. Закладка печінки відбувається приблизно на 3-4-му тижні ембріонального періоду, на 5-му тижні вона стає центром кровотворення. В печінці кровотворення відбувається екстраваскулярно за ходом капілярів, які вростають разом із мезенхімою всередину печінкових часток. Джерелом кровотворення у печінці є стовбурові клітини, які мігрували з жовткового мішка. З них утворюються бласти, які диференціюються у вторинні еритроцити. Водночас у печінці утворюються зернисті лейкоцити, головним чином нейтрофільні та еозинофільні. Окрім гранулоцитів, у печінці формуються гігантські клітини — мегакаріоцити. Кровотворення у печінці припиняється до кінця внутрішньоутробного періоду.
Кровотворення в тимусі. Закладка тимуса відбувається наприкінці 1-го місяця ембріонального періоду, на 7-8-му тижні епітелій тимуса починає заселятися стовбуровими клітинами крові, які диференціюються в лімфоцити тимуса. Збільшуючись кількісно, вони дають початок Т-лімфоцитам, які заселяють Т-зони периферійних органів кровотворення.
Кровотворення у селезінці. Селезінка закладається наприкінці 1-го місяця ембріонального розвитку. Із стовбурових клітин, які потрапляють до неї, відбувається екстраваскулярне утворення всіх видів формених елементів, тобто селезінка в ембріональному періоді є універсальним кровотворним органом. Утворення еритроцитів і гранулоцитів у селезінці досягає максимуму на 5-му місяці ембріонального розвитку, після чого в ній починає переважати лімфоцитопоез.
Кровотворення в лімфатичних вузлах. Перші Лімфатичні вузли закладаються на 7-8-му тижні ембріогенезу, а основна їх маса розвивається на 9-10-му тижні. Водночас відбувається проникнення в лімфатичні вузли стовбурових кровотворних клітин, з яких на ранніх етапах розвитку диференціюються еритроцити, гранулоцити, мегакаріоцити. Перші лімфоцити з’являються протягом 8-15-го тижнів. Починаючи з 16-го тижня, в лімфатичні вузли масово надходять попередники Т- і В- лімфоцитів, що пригнічує диференціацію інших формених елементів крові.
Із клітин-попередників диференціюються лімфобласти (великі лімфоцити), середні і малі лімфоцити. Диференціація Т- і В-лімфоцитів відбувається у Т- і В-залежних зонах лімфатичних вузлів.
Кровотворення в кістковому мозку. На 2-му місяці ембріонального розвитку відбувається закладка кісткового мозку, на 12-му тижні з’являються перші гемопоетичні елементи. Основна маса гемопоетичних елементів складається з еритробластів і попередників гранулоцитів. Стовбурові клітини кісткового мозку дають початок розвитку всім форменим елементам крові.
Постембріональний гемоцитопоез
Утворення і розвиток еритроцитів (еритроцитопоез), тромбоцитів (тромбоцитопоез), гранулоцитів (гранулоцитопоез), моноцитів (моноцитопоез) відбувається в гемопоетичній тканині червоного кісткового мозку (мієлоїдна тканина). Цей процес має назву мієлопоез.
Утворення та розвиток лімфоцитів і плазмоцитів відбуваються в кровотворній тканині, розташованій у селезінці, тимусі та лімфатичних вузлах (лімфоїдна тканина). Тому процес утворення цих клітин має назву лімфопоез.
Донині існувало безліч теорій кровотворення, які базувались на виділенні однієї або кількох родоначальних клітин, з яких утворюються всі види зрілих формених елементів. Також існували поліфілетичні теорії кровотворення, згідно з якими існує дві (дуалістична), три (тріалістична) та більше похідних клітинних форм. Сьогодні вони мають тільки історичне значення.
Нині загальновизнаною є унітарна теорія кровотворення, згідно з якою всі зрілі формені елементи крові є похідними однієї загальної клітини — родоначальниці.
Основні положення цієї теорії було сформульовано на початку XX ст. видатним російським гістологом О. О. Максимовим.
Вчений вважав, що існує спільна Клітина — родоначальниця всіх зрілих формених елементів крові, що має морфологію малого лімфоцита.
Сьогодні всі ці припущення підтверджено численними експериментами, які базуються на нових методах дослідження і дають можливість отримувати клітинні колонії або кровотворні колонії в селезінці смертельно опромінених мишей. Результати цих досліджень стали основою сучасної унітарної теорії кровотворення, згідно з якою всі зрілі формені елементи крові є похідними єдиної клітини, яка здобула назву стовбурової кровотворної клітини.
У дорослих ссавців стовбурові кровотворні клітини переважно зосереджені в червоному кістковому мозку, їхня кількість — близько 50 клітин на 105 ядерних клітин, у селезінці — 3,5 клітини, серед лейкоцитів крові — 1,4 клітини. Загальна кількість стовбурових кровотворних клітин у людини дорівнює близько 5∙1010. Дослідження чистої фракції стовбурових клітин за допомогою електронного мікроскопа довело, що їхня ультраструктура нагадує ультраструктуру малого темного лімфоцита. Стовбурова клітина має округлу чи овальну форму, діаметром близько 8 мкм, ядерно-цитоплазматична рівновага зсунута в бік ядра. Ядро діаметром близько 5 мкм, овальної чи округлої форми, з щільними скупченнями хроматину під ядерною мембраною (маргінальний шар). Цитоплазма має вигляд тонкого обідка, містить багато поодиноких рибосом, невелику кількість мітохондрій, гранулярну ендоплазматичну сітку.
Головними ознаками стовбурової кровотворної клітини є такі:
— поліпотентність, тобто здатність до диференціації у напрямку всіх видів формених елементів;
— здатність до самопідтримки протягом часу, близького до терміну існування організму людини;
— кількість мітозів, які може здійснити одна клітина, може перевищувати 100;
— стовбурова клітина у фізіологічних умовах ділиться дуже рідко, перебуваючи в G0-періоді клітинного циклу, незважаючи на високу здатність до проліферації;
— під дією несприятливих факторів (іонізуюче випромінювання, НВЧ-поле тощо) вона може дуже швидко розпочати поділ;
— стовбурові кровотворні клітини перебувають у стані постійної та інтенсивної репопуляції, тобто мігрують із одних кровотворних органів в інші.
Відповідно до сучасних уявлень про кровотворення, в усіх гістогенетичних рядах, які закінчуються утворенням зрілих формених елементів крові, виділяють такі класи клітин:
I клас — поліпотентні клітини-попередники, представлені стовбуровими кровотворними клітинами;
II клас — частково детерміновані клітини-попередники — напівстовбурові кровотворні клітини, потенція яких частково обмежена щодо подальшої їх диференціації, тобто з них можуть розвиватися не всі види формених елементів;
III клас — уніполярні клітини-попередники, здатні розвиватися тільки в одному напрямку під впливом певного виду гемопоетинів;
IV клас — морфологічно ідентифіковані проліферуючі клітини-попередники;
V клас — дозріваючі клітини, які втрачають здатність до мітотичного ділення;
VI — зрілі клітини, здатні до виходу в кров.
Еритроцитопоез (erythropoesis) — процес утворення та дозрівання еритроцитів, який відбувається в червоному кістковому мозку.
Джерелом розвитку еритроцитів є стовбурова кровотворна клітина (I клас), яка під впливом специфічного мікрооточення строми кісткового мозку в процесі поділу диференціюється в клітину-попередник мієлопоезу (II клас). Цю клітину ще називають колонієутворювальною одиницею гранулоцитів, еритроцитів, моноцитів, мегакаріоцитів (напівстовбурова кровотворна клітина), скорочено КУО—ГЕММ.
З цієї клітини утворюються більш детерміновані попередники двох видів:
— колонієутворювальна одиниця нейтрофільних гранулоцитів та еритроцитів (КУО—ГнЕ);
— колонієутворювальна одиниця мегакаріоцитів та еритроцитів (КУО—МгцЕ).
На наступному етапі відбувається утворення уніпотентної клітини-попередника — колонієутворювальної клітини еритроцитів (КУО—Е). Цей процес може відбуватися двома шляхами: з КУО—ГнЕ або з КУО—МгцЕ. Уніпотентну клітину еритропоезу ще називають еритропоетинчутливою клітиною, оскільки її подальша диференціація індукується еритропоети- ном, який виробляється у нирках. Еритропоетин посилює проліферацію еритропоетинчутливої клітини і її перетворення на проеритробласт, а також стимулює розвиток і розмноження еритроїдних клітин наступних стадій (рис. 58).
Проеритробласти — клітини IV класу еритроїдного ряду, округлої форми, діаметром 15-20 мкм.
Ядро велике, округлої форми, розташоване у центральній частині, має дрібносітчасту зернисту структуру, містить 1-3 ядерця.
Цитоплазма забарвлюється базофільно, містить багато рибосом, невелику центросому з двома центріолями, зерна феритину. Ядро оточене світлою перинуклеарною зоною. Проеритробласти діляться і перетворюються на базофільні еритробласти.
Базофільні еритробласти — клітини розміром 10-18 мкм.
Ядро містить Хроматин, розміщений променеподібно (як спиці в колесі).
Цитоплазма має інтенсивне базофільне забарвлення внаслідок великої кількості РНК. У цих клітинах розпочинається синтез гемоглобіну.
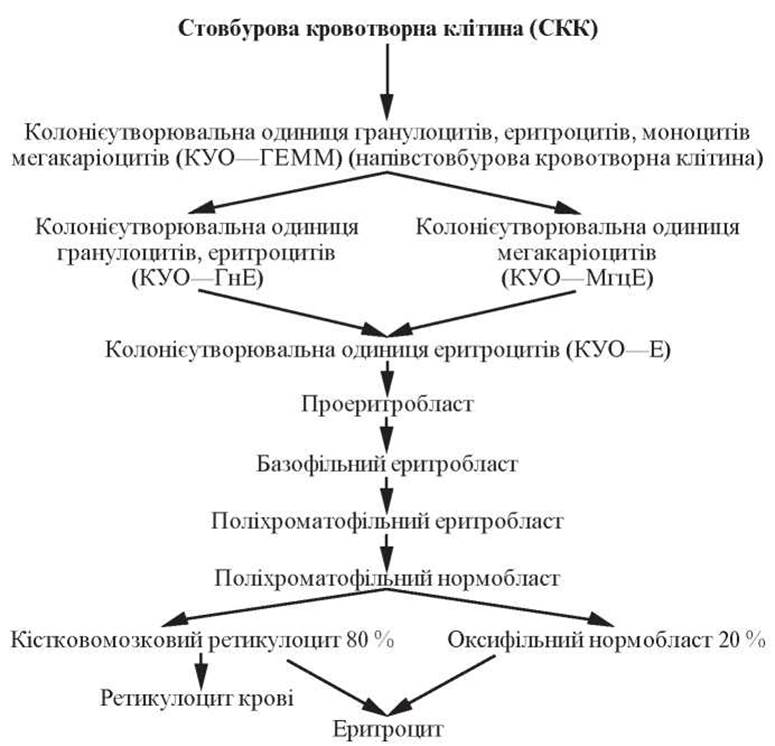
Рис. 58. Еритроцитопоез
Ці клітини діляться шляхом мітозу і, накопичивши певну кількість гемоглобіну, перетворюються на поліхроматофільні еритробласти.
Поліхроматофільні еритробласти — клітини розміром 1014 мкм.
Ядро менших розмірів, щільне, з чіткою колесоподібною структурою хроматину. Ядерця не визначаються.
Цитоплазма забарвлюється поліхромно, тобто і кислим, і основним барвниками. Оксифілія зумовлена наявністю гемоглобіну, базофілія — РНК. Гемоглобін у цих клітинах може розташовуватися дифузно (вся цитоплазма забарвлюється в сіруватий колір), плямами або у вигляді обідка навколо ядра. Рибосом менше, феритин розташовується агрегатами. Ці клітини діляться мітозом і перетворюються на поліхроматофільні нормобласти.
Поліхроматофільні нормобласти — клітини розміром до 10 мкм.
Ядро ущільнюється, втрачає колесоподібне розміщення хроматину, стає пікнотичним і майже безструктурним.
Клітини втрачають здатність до ділення, 80 % з них втрачають ядро, перетворюються на кістковомозкові ретикулоцити, які продовжують накопичувати гемоглобін і протягом 36-44 год дозрівають у кістковому мозку, після чого у вигляді еритроцитів надходять у кров. Частина кістковомозкових ретикулоцитів залишає Кістковий мозок, будучи не повністю насиченими гемоглобіном (це і є ретикулоцити крові). Решта 20 % поліхроматофільних нормобластів продовжують накопичувати гемоглобін, перетворюючись на оксифільні нормобласти, які втрачають ядро (шляхом виштовхування його з клітини або відриву від клітини ядерного фрагмента) і перетворюються на еритроцити.
Процес диференціації клітинних елементів еритропоезу відбувається протягом 100-140 год.
Клітини I, II і III класів у процесі їх ділення під час еритропоезу здійснюють 10-15 мітозів, а морфологічно визначені клітини еритроїдного класу — 5-6 мітозів. Із кожного проеритробласта утворюється 30-60 еритроцитів, процес їхнього утворення триває 6-8 днів. Загальна кількість еритроїдних клітин у кістковому мозку становить 3-1011.
Гранулоцитопоез (granulocytopoesis) — це процес утворення і розвитку гранулоцитів, що відбувається в червоному кістковому мозку.
Початковою клітиною гранулоцитопоезу є стовбурова кровотворна клітина червоного кісткового мозку і клітина-попередник мієлопоезу, які аналогічні описаним вище у розвитку еритроцитів. Це клітини I та II класу.
Наступним етапом є утворення більш детермінованої клітини-попередника гранулоцитів і моноцитів-макрофагів, колонієутворювальної одиниці гранулоцитів і моноцитів (КУО— ГМ). Із цієї клітини в процесі розвитку гранулоцитів утворюються уніпотентні попередники (III клас):
— базофілів — колонієутворювальні одиниці базофілів (КУО—Б);
— еозинофілів — колонієутворювальні одиниці еозинофілів (КУО—Ео);
— нейтрофілів — колонієутворювальні одиниці нейтрофілів (КУО—Нт).
Окрім перерахованого, уніпотентна клітина-попередник нейтрофілів може утворюватися також із колонієутворювальної одиниці нейтрофілів і еритроцитів (КУО—НтЕ).
Подальшу диференціацію клітин цього ряду стимулює гормон гранулопоетин (рис. 59).
Мієлобласти (myeloblastus) — великі клітини (до 20 мкм), що мають кругле ядро, розташоване в центрі, яке займає більшу частину клітини.
Ядро ніжно-сітчастої структури, на відміну від еритробластів, не має зерен хроматину, містить від 2 до 5 ядерець.
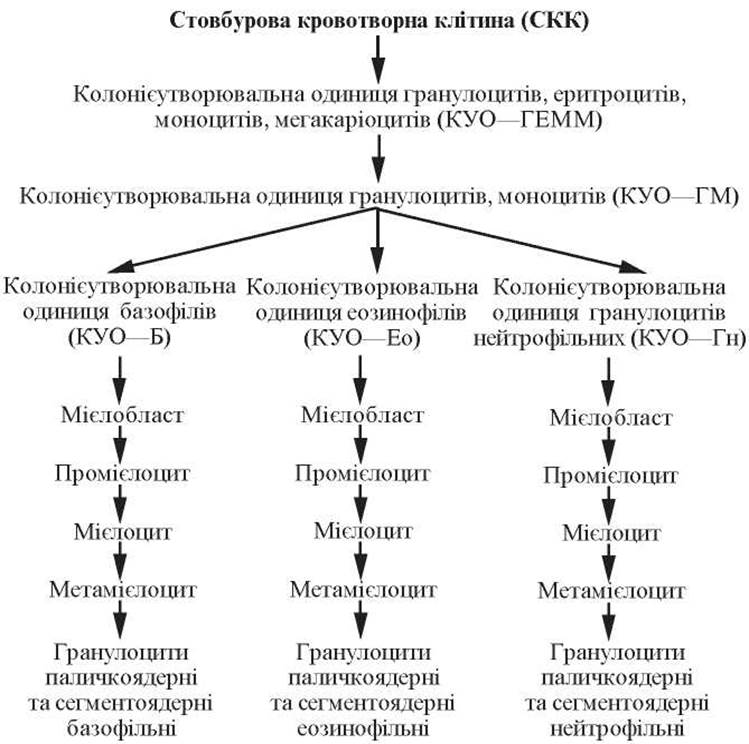
Рис. 59. Гранулоцитопоез
Цитоплазма мієлобластів базофільна, містить багато рибосом, мітохондрій, можна виявити неспецифічну азурофільну зернистість. Поблизу ядра розташовується центросома.
Мієлобласти діляться шляхом мітозу, перетворюючись на промієлоцити.
Промієлоцити (promyelocytus) — клітини розміром 12-20 мкм.
Ядро овальної форми, хроматин конденсований на периферії.
Цитоплазма містить багато розширених канальців гранулярної та агранулярної ендоплазматичної сітки, збільшену кількість вакуолей, комплекс Гольджі, який формується, багато мітохондрій. Формується зернистість, залежно від її характеру розрізняють три типи цих клітин — базофільні, еозинофільні та нейтрофільні.
Промієлоцити здійснюють один мітоз, внаслідок чого перетворюються на мієлоцити.
Мієлоцити (myelocytus) — клітини розміром від 8,0 до 12,0 мкм (нейтрофільні, еозинофільні, базофільні).
Ядро округлої форми, розташоване ексцентрично, містить щільні хроматинові тяжі, які чергуються зі світлими ділянками, ядерець немає.
Цитоплазма слабобазофільна або слабоеозинофільна, в ній зростає кількість специфічних зерен, накопичуються специфічні зерна, є добре розвинутий комплекс Гольджі, велика кількість розширених канальців гранулярної ендоплазматичної сітки; мітохондрій мало.
Ці клітини діляться шляхом мітозу двічі, перетворюючись на метамієлоцити.
Метамієлоцити (metamyelocytus) — це клітини округлої форми діаметром близько 8,0 мкм.
Ядро бобоподібної форми, ядерно-цитоплазматичний індекс зсунутий у бік цитоплазми.
Цитоплазма містить велику кількість специфічних гранул, добре розвинені Органели.
Існує три види цих клітин — базофільні, еозинофільні та нейтрофільні. Метамієлоцити вже не діляться, тому належать до класу клітин, які дозрівають (V клас). Метамієлоцити можуть потрапляти до периферійної крові, тоді їх називають юними.
Шляхом зміни форми ядра з метамієлоцитів утворюються паличкоядерні та сегментоядерні гранулоцити.
Моноцитопоез (monocytopoesis) — це процес утворення й розвитку моноцитів.
Клітини-попередники моноцитів перших двох класів було описано вище. З клітини-попередника гранулоцитів і моноцитів, макрофагів утворюється уніпотентна клітина-попередник моноцитів, або колонієутворювальна одиниця моноцитів (КУО— М, III клас) (рис. 60).
Моноцитобласт (monocytoblastus) — це велика клітина розміром близько 22,0 мкм, яка містить кругле ядро з вузьким обідком цитоплазми.
Ця клітина в процесі ділення диференціюється в промоноцит, який перетворюється на моноцит.
У процесі розвитку клітина зазнає таких змін: збільшується кількість цитоплазми, зменшується її базофілія, ядро набуває бобоподібної форми. Ядерно-цитоплазматичне співвідношення моноцита дорівнює 1:1.
Моноцити при цьому не є кінцевою стадією диференціації цього ряду, вони перетворюються в подальшому на макрофаги (гістіоцити — макрофаги) сполучної тканини.
На шляху від моноцитобласта до макрофага відбувається 7-8 мітозів.
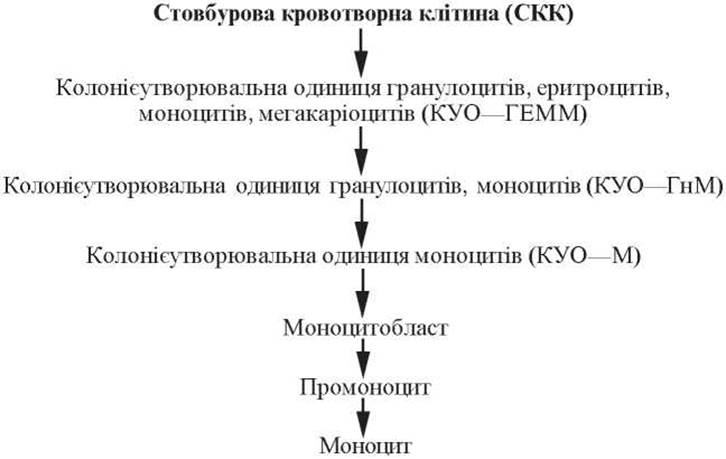
Рис. 60. Моноцитопоез
Тромбоцитопоез (trombocytopoesis) — це процес утворення та розвитку тромбоцитів, який складається з кількох послідовних стадій (рис. 61).
Уніпотентний попередник утворення тромбоцитів — це колонієутворювальна одиниця мегакаріоцитів (КУО—МГЦ), або тромбопоетинчутлива клітина.
Мегакаріобласт (megacaryoblastus) є наймолодшою морфологічно розрізнюваною клітиною тромбоцитопоезу, округлої форми, розміром 25,0-40,0 мкм.
Ядро містить рівномірно розподілений хроматин, 1-3 ядерця.
Цитоплазма базофільна, має дві зони: перинуклеарну, яка містить органели, та периферійну, пронизану угнутостями плазмолеми, які утворюють складні демаркаційні трубочки.
З мегакаріобласта утворюється промегакаріоцит.
Промегакаріоцит (promegacaryocytus) — це клітина розміром 40-80 мкм.
Ядро велике, починається його сегментація та огрубіння структури.
Цитоплазма менш базофільна, містить азурофільну зернистість, трубочки демаркаційної системи з’являються не тільки на периферії, а й у середній зоні цитоплазми. Промегакаріоцит диференціюється в подальшому в мегакаріоцит.

Рис. 61. Тромбоцитопоез
Мегакаріоцит (megacaryocytus) — найбільша клітина червоного кісткового мозку, розміром від 50,0-70,0 до 100,0 мкм.
Ядро поліморфне, фрагментоване із заглибленнями та вирізами, структура його грубосітчаста, ядерець немає.
Цитоплазма базофільна, містить азурофільну зернистість. За ходом демаркаційних трубочок цитоплазма розбивається на невеликі фрагменти, які відокремлюються від клітини та перетворюються на кров’яні пластинки. З однієї клітини може утворитися 3-4 тис. тромбоцитів.
Характерною особливістю мегакаріоцитів є те, що вони поліплоїдні, причому кількість хромосомних наборів у них може сягати 32-64. Поліплоїдизація цих клітин полягає в тому, що на шляху їх утворення з мегакаріобластів не відбувається поділу, а здійснюється 4-5 ендомітозів, внаслідок чого збільшується об’єм ядра та цитоплазми.
Лімфоцитопоез (lymphocytopoesis) — процес утворення та розвитку лімфоцитів, який також складається з кількох послідовних етапів.
Відповідно до положень унітарної теорії кровотворення, джерелом розвитку лімфоцитів є стовбурова кровотворна клітина I класу (СКК), з якої утворюється клітина-попередник лімфопоезу II класу. У подальшому розвиток цієї клітини відбувається у двох напрямках у відповідності з двома різновидами лімфоцитів — Т- і В-. В обох випадках утворюються уніпотентні попередники, які через лімфобласти (Т і В) перетворюються на Т- і В-лімфоцити.
Особливістю цих процесів є те, що зрілі клітини не є кінцевими елементами, їх подальший розвиток залежить від наявності антигенів. Тоді вони переходять у бластні форми і починають поділ. Цей, так званий антигензалежний процес диференціації лімфоцитів, відбувається у периферійних кровотворних органах — селезінці та лімфатичних вузлах (рис. 62).
Уважно ознайомившись з ілюстраціями (рис. 63-65), ви поглибите свої знання з гемоцитопоезу.
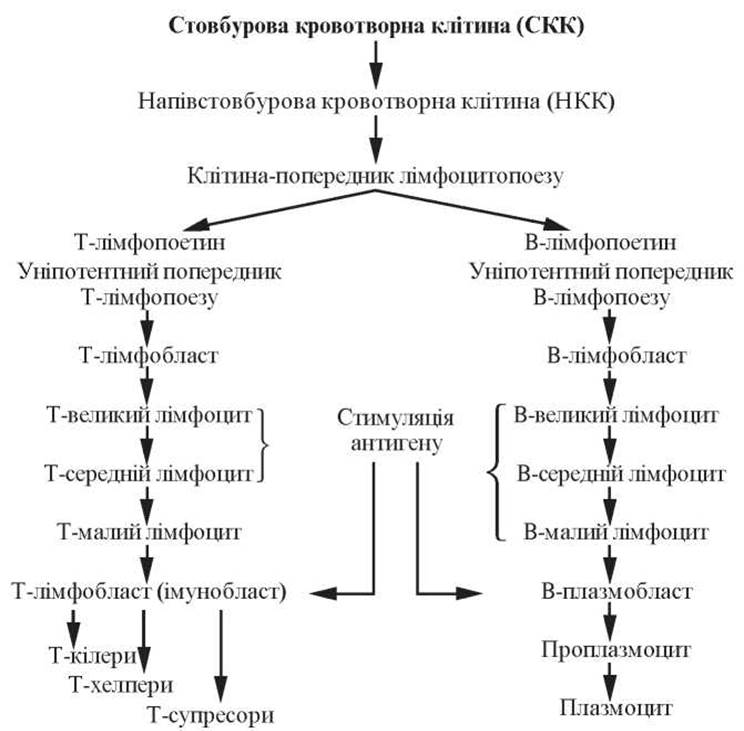
Рис. 62. Лімфоцитопоез та імуноцитопоез
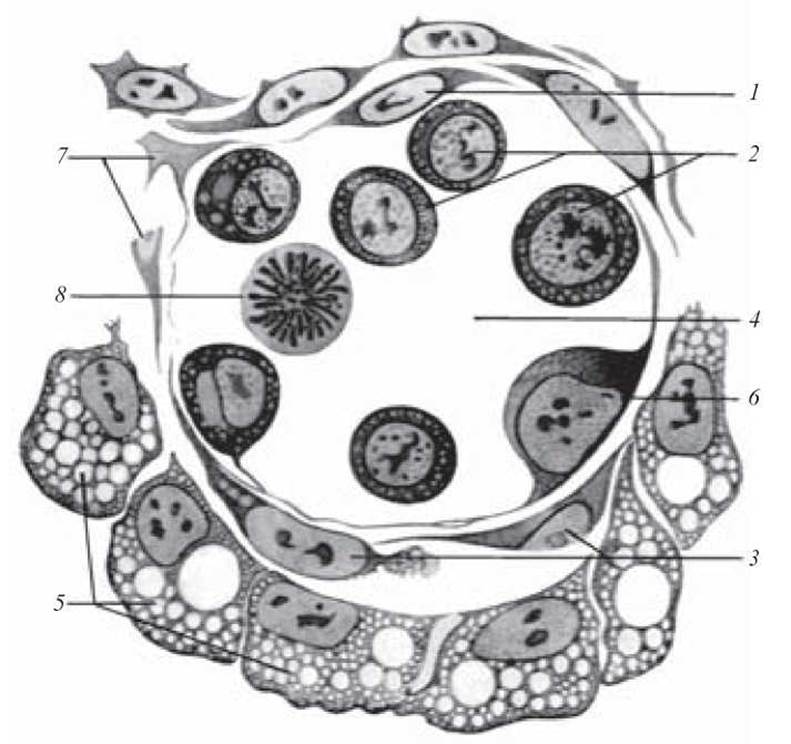
Рис. 63. Ембріональний гемоцитопоез. Поперечний зріз кров’яного острівця 8 1/2 -денного зародка кролика:
1, 6 — ендотелій; 2 — первинні кров’яні клітини; 3, 7 — мезенхімні клітини; 4 — просвіт судини; 5 — епітелій ентодерми; 8 — мітоз первинної кров’яної клітини
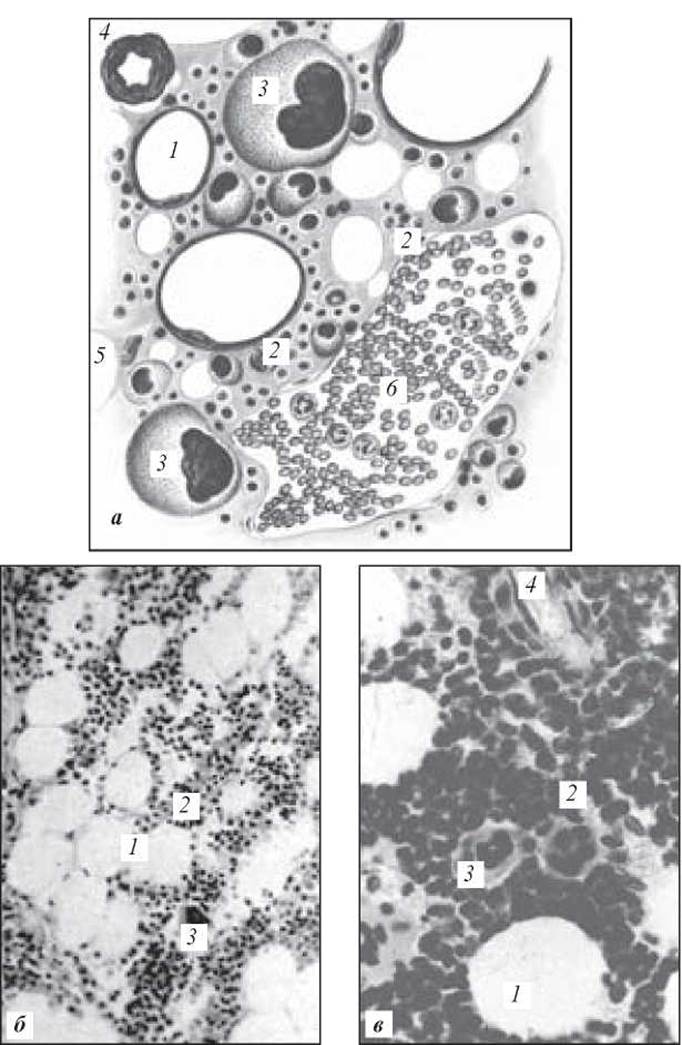
Рис. 64. Червоний кістковий мозок:
а — схема; б — забарвлення гематоксилін-еозином; в — фрагмент б; 1 — жирові клітини; 2 — острівці кровотворних клітин; 3 — мегакаріоцит; 4 — артерія; 5 — стромальна клітина; 6 — кровоносний синусоїдний капіляр (за О. В. Волковою)

Рис. 65. Еритробластичний острівець кісткового мозку миші. Електронна мікрофотограма. х 2376 (за Є. І. Заболотних). У безпосередньому зв’язку з макрофагом (М) перебуває пізній поліхроматофільний проеритроцит (П), який має нерівну поверхню та «пори». Видно довгий відросток ретикулярної клітини (показаний стрілкою)
Контрольні питання
1. Ембріональне кровотворення та його характеристика.
2. Постембріональне кровотворення.
3. Характеристика стовбурових, напівстовбурових та уніпотентних клітин.
4. Характеристика еритроцитопоезу.
5. Характеристика гранулоцитопоезу.
6. Характеристика моноцитопоезу.
7. Характеристика тромбоцитопоезу.
8. Характеристика лімфоцитопоезу.
Ситуаційні задачі
1. Якщо в базофільному еритробласті пригнітити біосинтез білка, який специфічний білок не буде утворюватися? Чи можлива подальша диференціація клітини?
2. При експериментальних дослідженнях із червоного кісткового мозку виділяли клітини еритробластичного ряду, в яких в основному закінчився синтез гемоглобіну. Які клітини виділено?
3. Якщо в ранньому неонатальному періоді пригнітити функцію тимуса, який вид гемопоезу порушується?
Приблизні екзаменаційні питання
1. Основні етапи ембріонального кровотворення.
2. Постембріональне кровотворення. Теорії кровотворення. Загальна схема.
3. Характеристика еритропоезу та тромбоцитопоезу.
4. Характеристика гранулоцитопоезу.
5. Лімфоцитопоез і моноцитопоез.
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.