Анатомия человека (с основами динамической и спортивной морфологии) - Иваницкий М. Ф. 2008
Введение в анатомию
Клетки и ткани
Органы тела образуются тканями, а последние — клетками. Их изучение составляет задачу двух морфологических наук — гистологии (науки о тканях) и цитологии (науки о клетке). Клетка — это живая саморетулируемая и самообновляемая системна, являющаяся основой строения, развития и жизнедеятельности всех животных и растительных организмов. В организме человека клетки различны по форме, величине, внутреннему строению и функциональному значению (рис. 1). Различают клетки шаровидной формы, веретенообразные, кубические, цилиндрические, звездчатые и др.
Величина клеток колеблется от 7 до 200 микрон (мкм).
Несмотря на многообразие форм, клетки имеют Общий план строения (рис. 2). Основными частями клетки являются Цитоплазма и Ядро.
Цитоплазма. Цитоплазма клетки неоднородна, в ней различают: цитолемму (плазмолемму), гиалоплазму, органеллы и цитоплазматические включения.
Цитолемма (плазмолемма) отделяет клетку от окружающей среды, регулирует Обмен веществ в клетке и обеспечивает постоянство ее внутренней среды. Толщина цитолеммы 9—10 нм*. В ее состав входят соединения белков и липидов с углеводами (Гликопротеиды и Гликолипиды), причем углевод содержащие части этих молекул, согласно А. Хэму и Д. Кормаку (1982), находятся в поверхностном слое цитолеммы — гликокаликсе. Холестерин, наоборот, располагается главным образом в глубоких ее слоях. На поверхности цитолеммы находятся так называемые рецепторы — молекулы, взаимодействующие с биологически активными веществами гормонами, медиаторами и т. и. и способствующие их «узнаванию» клеткой. Цитолемма участвует в поглощении клеткой крупных частиц (фагоцитозе) или макромолекул (пиноцитозе), подвергающихся процессам внутриклеточного пищеварения. Она обеспечивает выделение из клетки наружу ненужных продуктов обмена (экзоцитоз). По своему составу цитолемма близка мембранным цитоплазматическим органеллам, на долю которых вместе с цитолеммой приходится до 1/2 массы клетки.
Гиалоплазма — это основное вещество клетки, в котором располагаются все внутриклеточные образования, имеющие определенное Строение и выполняющие специфическую функцию. Различают органеллы мембранные и немембранные, общего значения и специальные. К органеллам общего значения относятся: эндоплазматическая сеть, Рибосомы, Митохондрии, Лизосомы, комплекс Гольджи (внутренний сетчатый аппарат) и центриоли.
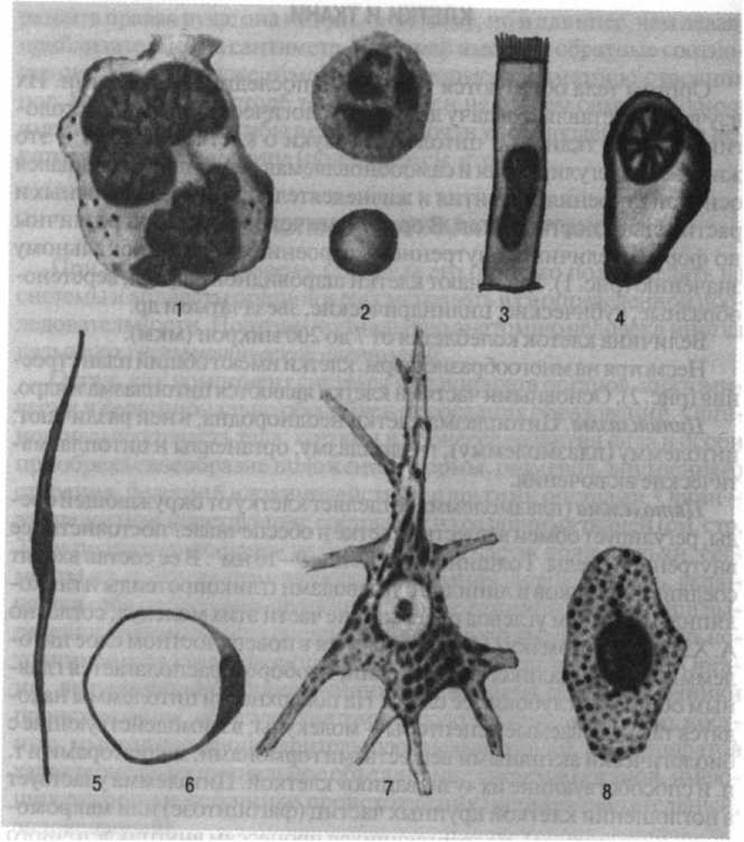
Рис. 1. Полиморфизм строения клеток:
1 - мегакариоцит костного мозга; 2 - нейтрофильный лейкоцит (вверху) и эритроцит (внизу): 3 - клетка мерцательного эпителия; 4 - плазматическая клетка; 5 - гладкомышечная клетка; 6 - жировая клетка; 7 - нейрон; 8 - гепариноцит
Эндоплазматическая сеть представляет собой систему канальцев, обеспечивающую транспорт веществ из окружающей среды и внутри клетки. Незернистая (гладкая) эндоплазматическая сеть участвует в синтезе углеводов и липидов, а зернистая, с рибосомами на поверхности, в синтезе белка.
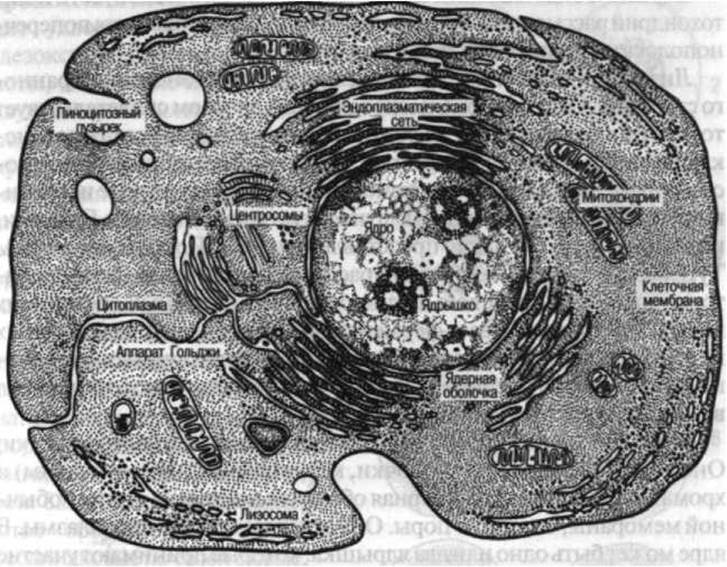
Рис. 2. Строение клетки (по Прокофьевой-Бельговской)
Рибосомы вырабатывают белок, причем специфический для каждого вида клеток.
Митохондрии содержат макроэргические соединения и являются источником энергии.
Лизосомы содержат большое количество ферментов и осуществляют внутриклеточное Пищеварение.
Комплекс Гольджи (внутренний сетчатый аппарат), состоящий из цистерн, канальцев и пузырьков, является местом скопления веществ, секретируемых клеткой.
Центриоли, образующие центросому, принимают участие в делении клетки.
Специальные органеллы связаны со специфической функцией клетки. К ним относятся миофибриллы в мышечных клетках, нейрофибриллы — в нервных, тонофибриллы — в эпителиальных.
Включения — это зернышки белка, капельки жира, пигмента, которые могут быть или не быть в клетке, в отличие от органелл — постоянных ее образований.
Строение эндоплазматической (саркоплазматической) сети и митохондрий рассмотрено более подробно на примере волокна поперечнополосатой мышечной ткани (см. стр. 35).
Лизосомы во многом отличаются от других органелл мембранного строения. О функциональной важности лизосом свидетельствует то, что они содержатся у млекопитающих в клетках всех типов, исключая лишь зрелые эритроциты. Структурная изменчивость лизосом определяется тем, что их мембраны могут иметь разные источники происхождения: эндоплазматическую сеть, комплекс Гольджи, цитолему и др. Согласно представлениям А. А. Покровского и В. А. Тутельяна, в мембранах лизосом располагаются ферментные рецепторы, проявляющие свою активность при аварийных состояниях клетки. Это обеспечивает ферментную защиту клеток от ненужных или чужеродных продуктов обмена веществ и патогенных микроорганизмов, на что впервые и задолго до открытия лизосом обратил внимание И.И. Мечников.
Ядро. Как и цитоплазма, ядро является основной частью клетки. Оно состоит из ядерной оболочки, кариоплазмы (нуклеоплазмы) и хроматиновых структур. Ядерная оболочка построена по типу обычной мембраны, содержит поры. Она отделяет ядро от цитоплазмы. В ядре может быть одно или два ядрышка, которые принимают участие в обмене веществ, в том числе и в образовании рибосом.
Кариоплазма, представляющая собой сетчатое образование, содержит ФЕРМЕНТЫ и другие химически активные вещества. В ней происходит Синтез белка.
Хроматиновые структуры в делящейся клетке образуют хромосомы — материальные Носители наследственной информации. Их 23 пары, одна из которых состоит из хромосом, называемых половыми — хромосомы X и хромосомы Y.
Клетки человеческого организма размножаются путем непрямого деления — митоза и прямого деления — амитоза. Для половых клеток характерен иной тип деления — Мейоз, проявляющийся в уменьшении генетического материала (в отличие от соматических клеток половые имеют не двойной, а одинарный набор хромосом. Считают, что митоз в отличие от амитоза характеризуется более высокой активностью ядра клетки. Процесс митоза, или кариокинеза (рис. 3), на основании морфологически видимых изменений условно подразделяют на четыре фазы: профазу, метафазу, анафазу и телофазу.
Их общая продолжительность составляет от одного до полутора часов. Период между клеточными делениями называется интерфазой. Она подразделяется на три периода длительностью около 19часов, на протяжении которых происходит удвоение молекул ДНК (дезоксирибонуклеиновой кислоты) для обеспечения генами двух дочерних клеток. Клетка, подвергающаяся делению непрерывно цикл за циклом, не может выполнять свои специализированные Функции. Для реализации их ей приходится на время или навсегда прекратить митотические деления.
1. Профаза. Изменения клетки в процессе митоза начинаются с того, что ядро набухает, хроматиновое вещество приобретает форму сплошной, спутанной нити, образующей рыхлый клубок. Затем в ней четко выявляются обособленные участки, неодинаковые по своей форме, — хромосомы (в менее различимом виде они присутствуют и в интерфазном ядре), каждая из которых подразделяется на две дочерние хромосомы. Оболочка ядра растворяется. Ядрышко исчезает. Центросома делится на две центриоли, расходящиеся к двум противоположным полюсам клетки.
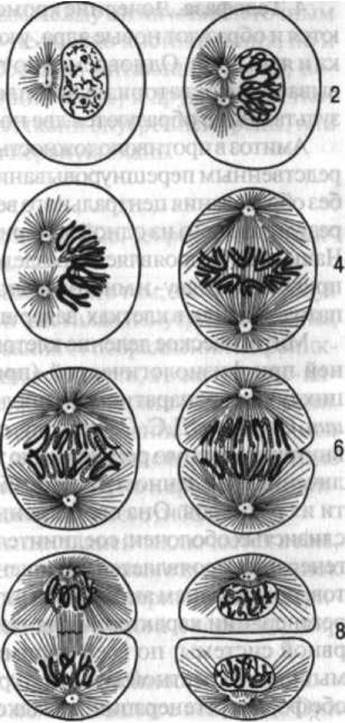
Рис. 3. Непрямое деление (митоз):
1 — клетка в покое; 2 профаза (ранняя стадия); 3 — профаза (поздняя стадия); 4 — метафаза; 5 — анафаза; 6 — телофаза; 7—8 — Разделение цитоплазмы и ядра
2. Метафаза. К этому времени между центриолями, занимающими полюсные положения в клетке, образуется веретено из ахроматиновых нитей, принимающих участие в передвижении хромосом. Хромосомы располагаются в центральной, экваториальной, плоскости клетки, образуя фигуру, напоминающую звезду. Они становятся короче и толще; еще более заметным становится подразделение каждой из них на две дочерние.
3. Анафаза. Дочерние хромосомы расходятся к полюсам клетки и собираются у центриолей, образуя фигуру двойной звезды.
4. Телофаза. Дочерние хромосомы собираются вместе, уплотняются и образуют новые ядра, у которых появляются ядерная оболочка и ядрышки. Одновременно тело клетки постепенно перешнуровывается в экваториальной плоскости и разделяется надвое, в результате чего образуются две новые клетки.
Амитоз в противоположность кариокинезу характеризуется непосредственным перешнуровыванием ядра и цитоплазмы (совершается без образования центрального веретена и спирализации хромосом), в результате чего из одной материнской клетки возникают две новые. Чаще амитоз проявляется разделением ядра, но не клетки в целом, что приводит к ее дву- и многоядерности. Он наблюдается в основном при патологии или в клетках, завершающих свой жизненный цикл.
Митотическое деление клеток обеспечивает самообновление тканей при физиологической (присущей нормальному жизненному циклу) или репаративной (после повреждения тканей) их регенерации. Согласно Д. С. Саркисову, следует разграничивать клеточную и внутриклеточную регенерацию тканей. Первая характеризуется увеличением численности клеток (благодаря делению) при неизменности их размеров. Она свойственна эпителиальному покрову кожи и слизистых оболочек, соединительным тканям. Внутриклеточная регенерация проявляется увеличением размера клеток и их компонентов, повышением активности внутриклеточных структур. Эта форма регенерации характерна для ганглиозных клеток центральной нервной системы; по всей видимости, она преобладает в сердечной мышце и скелетной мускулатуре. Во многих случаях проявляются обе формы регенерации; в гладкой мускулатуре, печени, почках и др.
Наряду с клеточными формами организации живого вещества существуют и неклеточные структуры. К ним можно отнести межклеточное основное вещество. Клеточные структуры могут иметь упрощенное (эритроциты, кровяные пластинки) или усложненное (поперечнополосатое мышечное волокно, нейрон) строение. По мнению А.Н. Студитекого, появлению клеток в живой природе предшествуют филогенетически более древние доядерные структуры (прокариоты), из которых клетки сформировались в процессе эволюции живой материи.
Ткани
Ткань — это исторически сложившаяся система клеток и неклеточных структур, обладающих общностью строения и происхождения, специализированная на выполнении определенных функций.
В строении каждого органа принимает участие не одна какая-либо ткань, а различные виды тканей. Например, в строении костной ткани — костные клетки с расположенным между ними межклеточным веществом, а в образовании кости — не только костная ткань, но и мышечная (в стенке кровеносных сосудов, питающих кость), нервная (в образованиях, иннервирующих кость) и другие виды тканей. Различают ткани эпителиальные, ткани внутренней среды, или соединительные, Мышечные ткани и нервную ткань.
Эпителиальные ткани
Эпителиальные ткани выполняют защитные, секреторные, экскреторные и всасывательные функции.
Защитная функция этих тканей заключается в том, что они, образуя наружный слой кожи, ее эпидермис, и выстилая изнутри все органы, предохраняют глубже лежащие образования от повреждений и проникновения в них микробов и других вредных веществ, а в желудочно-кишечном тракте — от разрушения его стенки пищеварительными соками. Через эпителиальные ткани происходит всасывание переваренной пищи в желудочно-кишечном канале, а также выведение из организма продуктов обмена веществ, не только ненужных, но и вредных (например, мочи, задержка которой в организме ведет к его гибели).
Секреторная функция эпителиальных тканей заключается в том, что они, участвуя в образовании желез, их концевых отделов и стенок выводных протоков, вырабатывают секреты, например пищеварительные соки (железы органов пищеварительной системы), жир (сальные железы), пот (потовые железы).
Все эпителиальные ткани имеют общие особенности строения.
В них незначительно количество межклеточного вещества. Они образуют пласты из клеток, плотно прилегающих друг к другу и расположенных на базальной мембране. Эпителиальные ткани обладают высокими регенеративными свойствами, в них всегда сохраняются клетки, способные к митозу.
Источниками происхождения эпителиальных тканей являются внутренний (энтодермальный), средний (мезодермальный) и наружный (эктодермальный) зародышевые листки (см. стр. 44).
Классификация эпителиальных тканей основана на особенностях их строения и функции, а также на участии в образовании тех или иных органов (рис. 4).
По количеству слоев клеток различают эпителий однослойный (например, выстилающий изнутри кишечник) и многослойный (например, кожный покров или покрывающий спереди роговую оболочку глаза).
По форме клеток выделяют плоский, кубический, цилиндрический (и его разновидность —мерцательный) и переходный эпителий. Плоский эпителий покрывает серозные оболочки (плевру, брюшину), кубический участвует в образовании стенки канальцев Почки, цилиндрический выстилает слизистую оболочку органов пищеварения (Желудок, кишки), мерцательный - дыхательные пути, маточные трубы, а переходный - пути выведения мочи (почечные лоханки, Мочеточники, Мочевой пузырь).
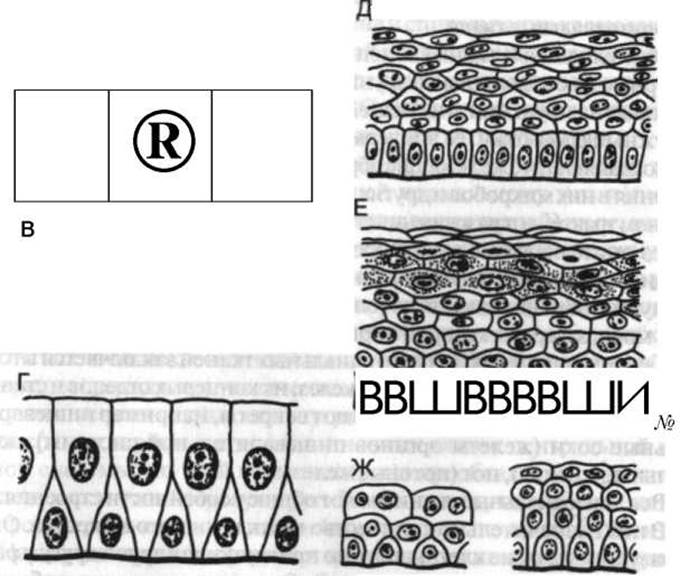
Рис. 4. Схема строения различных эпителиев:
А, Б, В - однослойные (А - плоский, Б - кубический, В - цилиндрический); Г - многорядный; Д, Е - многослойные (Д - неороговевающий, Е - ороговевающий); Ж - переходный: слева — в состоянии наполнении органа, справа — в его спавшемся состоянии (по Е. А. Шубниковой, 1981)
Многослойный эпителий (обычно плоский) имеет две разновидности: ороговевающий -выстилающий поверхность кожи и неороговевающий — выстилающий с внутренней поверхности слизистую оболочку полости рта, глотки, начального отдела пищевода, прямой кишки, влагалища, роговицу глаза.
Железистый эпителий выделяет свои продукты на поверхность эпителиальной выстилки в полые органы или непосредственно в Кровь и лимфу, т. е. образует железы (органы) внешней и внутренней секреции. Железы бывают одноклеточные (например, бокаловидные клетки кишечника, выделяющие слизь) и Многоклеточные, которые образуются путем выпячивания слоя эпителия. Они имеют разнообразные форму и функцию. Различают железы альвеолярные, трубчатые и смешанные, т. е. альвеолярно-трубчатые. Кроме того, железы могут быть простыми, разветвленными и сложными.
Простые трубчатые железы представляют собой трубкообразные углубления (например, кишечные железы).
Разветвленные трубчатые железы состоят из нескольких простых, открывающихся в общий выводной проток (например, железы привратниковой части желудка, железы матки).
Сложные трубчатые железы образованы несколькими разветвленными трубчатыми железами, открывающимися в общий выводной проток (например, слезные железы, Печень, яички).
Простые альвеолярные железы имеют вид небольших пузырьков или мешочков (например, сальные железы).
Разветвленные альвеолярные железы состоят из нескольких простых альвеолярных желез, открывающихся в общий выводной проток (например, железы век).
Сложные альвеолярные железы образуются из нескольких разветвленных альвеолярных желез, имеющих общий выводной проток (например, поджелудочная, молочная железы).
Простые альвеолярно-трубчатые железы представляют собой трубки, имеющие на своем конце расширения (встречаются, например, в привратниковой части желудка).
Разветвленные альвеолярно-трубчатые железы состоят из нескольких простых альвеолярно-трубчатых желез (например, железы мочеиспускательного канала).
Сложные альвеолярно-трубчатые железы построены из разветвленных альвеолярно-трубчатых желез (например, Легкие, Предстательная железа). В некоторых случаях описанные железы имеют усложнения в строении. Так, простые трубчатые железы могут иметь на своем конце завитки (например, потовые железы).
Составными частями многоклеточной железы являются: дно, тело, шейка, устье, а у более крупных желез, кроме того, хорошо выраженный выводной проток.
Расширенное и слепо заканчивающееся место железы — ее тело и дно — выстлано секреторным эпителием. В этом участке железы осуществляется процесс секреции. Образование секрета в разных железах происходит различным путем: в одной группе желез без нарушения клеток (например, бокаловидных), в другой (например, молочной железе) связано с частичной гибелью клеток, в третьей (например, сальных железах) сопровождается разрушением клеток. Разрушенные клетки замещаются новыми, которые образуются путем размножения более глубоко находящихся клеток, прилегающих к базальной мембране и способных к делению (размножению).
В отличие от желез внешней секреции эндокринные железы лишены выводных протоков (см. стр.409).
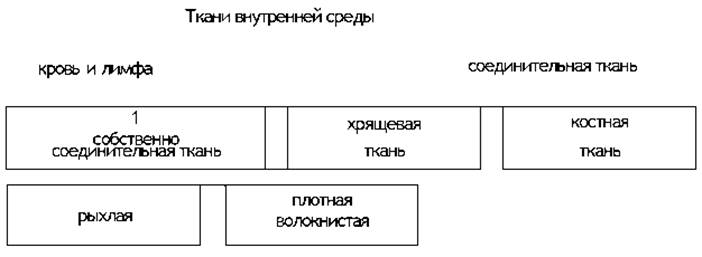
Ткани внутренней среды
Ткани внутренней среды, или Соединительные ткани, отличаются значительным развитием межклеточного вещества. Они подразделяются на ткани с преобладанием трофических, опорных или защитных функций.
В основном трофическую, т. е. питательную, функцию выполняют кровь, лимфа, отчасти рыхлая Соединительная ткань, которые участвуют в снабжении организма питательными веществами и кислородом, а также в удалении продуктов обмена веществ и углекислоты.
Эти ткани выполняют также защитную функцию, которая заключается в выработке веществ, разрушающе действующих на попавшие в организм микробы, в продуцировании антитоксинов и иммунных тел, в способности некоторых клеток поглощать и переваривать микробы и инородные вещества (фагоцитарная способность).
Опорную функцию выполняют главным образом плотная соединительная ткань, Хрящ, кость и отчасти рыхлая соединительная ткань. Отдельные виды тканей внутренней среды участвуют в образовании твердого и мягкого скелетов. Это не только кости и их соединения, но также связки, фасции, межкостные и межмышечные перепонки, каркас внутренних органов и пр.
Существует ряд классификаций тканей, в частности разграничивающая соединительную ткань, кровь и лимфу как самостоятельные типы тканей. Наиболее распространенной является следующая классификация (см. схему 1).
Кровь и лимфа. Это жидкие виды тканей внутренней среды. Они содержат около 80% воды и около 20% органических веществ, имея удельный вес около 1,055. Общее количество крови достигает у мужчин приблизительно 4,5 л, а у женщин — 4 л. Кровь и лимфа состоят из плазмы и форменных элементов (рис. 5). Общая масса форменных элементов составляет 1/3—2/5 объема всей крови, в то время как плазмы — 2/3—3/5 этого объема. Среди форменных элементов крови различают красные и белые кровяные тельца (см. схему 2).
Схема 1.
У человека красные кровяные тельца, эритроциты, в отличие от белых, лишены ядер. Они имеют форму дисков с плоским углублением на обеих сторонах. В 1 мм2 крови содержится в среднем 4,5-5 млн. эритроцитов. Общая поверхность эритроцитов составляет примерно 3200 м2, причем во время каждого вдоха в обмене газов в легких участвует приблизительно 250 м2 их общей поверхности. В состав эритроцитов входит красящее вещество крови - Гемоглобин, составляющий приблизительно 1/3 их общей массы. Он обладает свойством соединяться с кислородом, играя тем самым важную роль в газообмене. Кровь при этом меняет вишнево-красный цвет на светло-красный. В составе крови взрослого человека около 660 г гемоглобина. Место размножения эритроцитов - красный Костный мозг. Продолжительность их жизни — 80-120 дней.
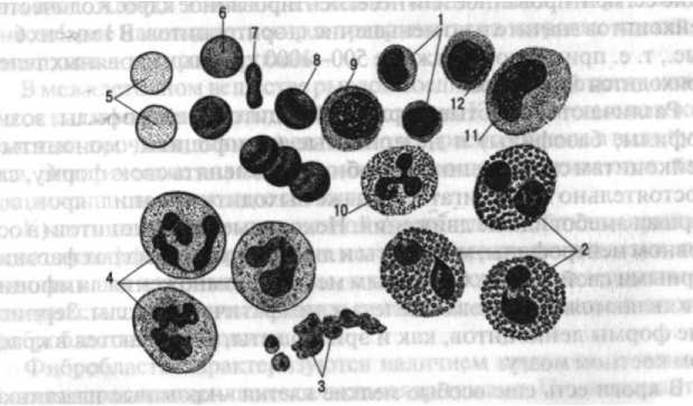
Рис. 5. Кровь человека. Общий вид эритроцитов, лейкоцитов и тромбоцитов в окрашенном препарате (увелич. в 1 000 раз):
1 - малые лимфоциты; 2 - эозинофильные лейкоциты; 3 - группа кровяных пластинок; 4 - нейтрофильные лейкоциты; 5-8 - эритроциты в разных положениях; 9 - большой лимфоцит; 10 - базофильный лейкоцит; 11 - моноцит; 12 - средний лимфоцит
Схема 2.
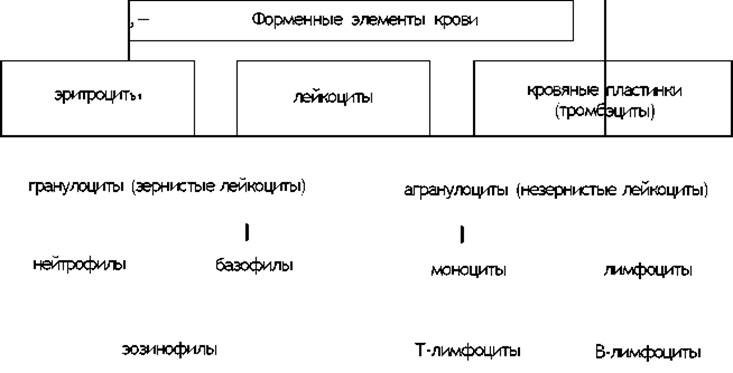
Белые кровяные тельца, лейкоциты, имеют в своей цитоплазме одно сегментированное или несегментированное ядро. Количество лейкоцитов значительно меньше числа эритроцитов. В 1 мм5 их 6—8 тыс., т. е. примерно на каждые 500-1000 красных кровяных телец приходится одно белое.
Различают зернистые формы лейкоцитов (нейтрофилы, эозинофилы, базофилы) и незернистые (лимфоциты, моноциты). Лейкоцитам свойственна способность изменять свою форму, самостоятельно передвигаться и даже выходить из капилляров, совершая амебовидные движения. Некоторые из лейкоцитов (в основном нейтрофилы, моноциты и лимфоциты) обладают фагоцитарными свойствами. Основным местом размножения лимфоцитов являются вилочковая железа и Лимфатические узлы. Зернистые формы лейкоцитов, как и эритроциты, развиваются в красном костном мозгу.
В крови есть еще особые мелкие клетки — кровяные пластинки (тромбоциты) (размером 2-5 мкм, т. е. в 2-4 раза меньше красных и в 5—7 раз меньше белых кровяных телец. Они принимают участие в свертывании крови. В 1 мм3 крови примерно 200-300 тыс. тромбоцитов.
Лимфа, как и кровь, состоит из лимфоплазмы и форменных элементов. По своему составу лимфоплазма сходна с плазмой крови, но содержит меньше белков. Из форменных элементов в лимфе находятся преимущественно лимфоциты и моноциты (см. стр. 323).
Основу кроветворных органов костного мозга, лимфатических узлов, селезенки — образует ретикулярная ткань. Кроме того, она входит в состав слизистой оболочки желудочно-кишечного тракта, дыхательных путей и некоторых других органов. Клетки ретикулярной ткани образуют своеобразную сеть и могут в некоторых случаях высвобождаться из связи, становиться подвижными и самостоятельными. Они обладают фагоцитарными свойствами.
Фагоцитарные свойства присущи тесно связанным с ретикулярной тканью клеткам эндотелия капилляров, печени и других органов. Это послужило основанием для выделения так называемой ретикуло-эндотелиальной системы, объединяющей ретикулярную ткань, эндотелий некоторых капилляров, клетки рыхлой соединительной ткани, обладающие способностью к активному перемещению и фагоцитозу, а также моноциты и лимфоциты крови. Все названные образования выполняют в организме защитную функцию.
Рыхлая соединительная ткань. Она имеет в организме большое распространение: сопровождает Кровеносные сосуды и нервы на всем их протяжении, располагается между органами и в подкожном жировом слое. Клетки этого вида соединительной ткани могут в своей цитоплазме накапливать жир и превращаться в жировые клетки, а сама ткань — в жировую ткань.
В межклеточном веществе рыхлой соединительной ткани находятся коллагеновые и эластические волокна. Коллагеновые волокна толстые, прочные, плохо растягиваются, расположены в виде пучков. Эластические волокна - тонкие, хорошо растягиваются, возвращаясь вновь в исходное состояние.
К клеточным элементам рыхлой соединительной ткани принадлежат недифференцированные мезенхимные клетки и клетки, попавшие в нее из крови и лимфы, к которым относятся зернистые и незернистые лейкоциты. Основными клетками этой ткани являются фибробласты и гистиоциты.
Фибробласты характеризуются наличием крупного, овальной формы ядра с одним или несколькими ядрышками. Цитоплазма этих клеток может быть зернистой и гомогенной с наличием цитоплазматических отростков. Фибробласты встречаются в различных формах дифференцировки. К малодифференцированным элементам рыхлой соединительной ткани относятся молодые фибробласты и ретикулярные клетки, а кроме того, некоторые эндотелиальные клетки, отделяющие русло кровеносного сосуда от окружающей рыхлой соединительной ткани.
Гистиоциты имеют ядро, часто вдавленное с одной стороны, овальной или неправильной формы и меньших размеров, чему фибробластов. Они отличаются от фибробластов тем, что контуры их хорошо очерчены и нигде не сливаются с основным, межклеточным, веществом.
Кроме фибробластов и гистиоцитов в рыхлой соединительной ткани находятся и другие клетки (пигментные, тучные, жировые, плазматические, лимфоциты и лейкоциты).
Плотная волокнистая соединительная ткань. Она встречается виде фиброзной соединительной ткани кожи, сухожилий и в виде эластической соединительной ткани. Плотная оформленная фиброзная соединительная ткань кожи характеризуется значительной прочностью, что обусловливается хорошим развитием коллагеновых пучков, переплетающихся между собой, а также эластических волокон. В этой ткани имеются фиброциты, закончившие свое развитие, и гистиоциты.
У плотной оформленной фиброзной ткани сухожилий толстые пучки коллагеновых волокон идут параллельно. В фасциях они располагаются в различных направлениях. Между пучками находятся небольшие щели с лежащими в них фиброцитами, называемыми сухожильными клетками. Эта ткань образует сухожилия, фасции связки.
Плотная оформленная эластическая ткань имеет значительное количество эластических волокон, образующих без подразделения на пучки целые большие тяжи или пластинки. Из этой ткани построены желтые связки позвоночного столба.
Хрящевая ткань. В зависимости главным образом от характера межклеточного вещества хрящевая ткань встречается в виде гиалинового, волокнистого и эластического хряща.
Гиалиновый, или стекловидный, хрящ (рис. 6, А) в естественном состоянии имеет бесструктурное, однородное межклеточное вещество состоящее из хондрина, хондромукоида и хондроитинсерной кислоты Его клетки располагаются в небольших полостях и заполняют их целиком. Межклеточное вещество содержит тонкие соединительнотканные волокна. Этот вид хрящевой ткани широко распространен. Он покрывает суставные поверхности костей, находится в реберных хрящах, хрящах гортани (кроме надгортанника), трахеи и бронхов.
Волокнистый хрящ встречается в межпозвоночных и суставных дисках и представляет собой волокнистую соединительную ткань, имеющую хрящевые клетки, пропитанную, как и гиалиновый хрящ, хондрином.
Эластический хрящ (рис. 6, Б) характеризуется большим количеством эластических волокон в межклеточном веществе, благодаря чему он имеет желтоватый цвет, резко отличающий его по внешнему виду от гиалинового хряща. Эластический хрящ встречается главным образом в надгортаннике и ушной раковине.
Хрящи, особенно гиалиновый, могут подвергаться обызвествлению. Это - одна из их характерных возрастных особенностей.
В этом отношении на первом месте стоят хрящи гортани (прежде всего щитовидный) и реберные.

Рис. 6. Схема строения зрелого гиалинового (А) и эластического (Б) хрящей:
1 - надхрящница; 2 - хондробласты; 3 - хондроциты; 4 - межклеточное вещество хряща; 5 - эластические волокна (по Стеопов, из книги Е.А. Шубниковой)
Костная ткань. Характерными особенностями костной ткани являются прочность и упругость, благодаря которым она может выполнять свои опорные функции. Костная ткань состоит из клеток и межклеточного вещества, содержащего органические (оссеин и оссеомукоид) и неорганические (соли, главным образом кальция) соединения. Различают два вида костной ткани: грубоволокнистую и пластинчатую. Грубоволокнистая костная ткань у человека находится лишь в местах прикрепления к кости сухожилий и в области зарастающих швов черепа.
Костям человека свойственно в основном пластинчатое строение. Находящиеся в пластинках параллельные пучки коллагеновых волокон идут в определенных направлениях соответственно действующим силам. Они пропитаны неорганическими соединениями, что и обусловливает прочность этого вида костной ткани, уступающую только прочности эмали зуба. Клетки костной ткани — остеоциты — находятся в полостях межклеточного вещества. Эти полости соединяются между собой тонкими канальцами, по которым сообщаются и отростки остеоцитов, что придает всей структуре костной ткани синцитиальный характер.
Кроме остеоцитов в костной ткани имеются остеобласты и остеокласты. Первые образуют костную ткань, а вторые разрушают, способствуя ее непрерывному обновлению.
Изучение микроскопического строения кости показало, что кость состоит из остеонов и интерстициальных костных пластинок (рис. 7). Остеон является основной структурной единицей кости. Он представляет собой систему как бы вставленных друг в друга костных цилиндров в виде пластинок остеона. Система концентрически расположенных пластинок окружает несколькими слоями центральный канал остеона. Коллагеновые волокна имеют в различных слоях пластинок разное направление, что способствует механической прочности остеона. Между остеонами находятся интерстициальные костные пластинки. Снаружи слой остеонированного костного вещества ограничен непрерывным слоем наружных окружающих костных пластинок, а изнутри, со стороны костномозговой полости, — слоем внутренних окружающих костных пластинок.
В центральном канале остеона проходят кровеносные сосуды и нервы. Калибр костных каналов составляет от 1/50 до 1/5 мм, в зависимости от величины кости и от положения канала по отношению к ее наружной поверхности. Более крупные кости обладают более широкими костными каналами. Поверхностно лежащие каналы обычно шире тех, которые расположены глубоко.
Остеоны, интерстициальные и окружающие пластинки перестраиваются на протяжении жизни в связи с механическими условиями функционирования кости, возрастом, особенностями питания и другими факторами. Считается, что длительность жизни остеоцита достигает 25 лет. Для взрослого человека обновление компактного костного вещества составляет 2,5%, а губчатого -10% в год. В детском возрасте интенсивность обновления кости выше.
О механических свойствах соединительной ткани можно судить по ее крепости на растяжение, сжатие, разрыв, скручивание, изгиб. Данные такого рода составляют новый раздел морфологии, называемый биосопроматом.

Рис. 7. Микроскопическое строение остеонированной кости (по Хэму)
Как известно, прочность на растяжение выражается той наименьшей отнесенной к единице поперечного сечения (1 мм2) величиной нагрузки, от которой ткань разрывается или деформируется. Свежее компактное костное вещество выдерживает сопротивление на растяжение до 10-12 кг, реберный (гиалиновый) хрящ - 0,5 кг, сухожилия, т. е. плотная фиброзная ткань, — 7 кг.
В отношении сопротивления на сжатие кость в 10 раз крепче хряща. Крепость кости на сжатие (12-16 кг) раза в полтора больше крепости на растяжение. Крепость гиалинового хряща на сжатие (1,5 кг) в 3 раза больше крепости на растяжение. Свежая кость в 5 раз прочнее железобетона как на сжатие, так и на растяжение. Для раздробления бедренной кости давлением нужно приблизительно 3000 кг, для раздробления же большеберцовой кости - не менее 4000 кг.
Сопротивление на изгиб при определенном весе кости достигает наибольшей величины в том случае, когда внутренний диаметр трубчатой кости относится к наружному примерно как 8:11. Улиц молодого возраста эта сопротивляемость больше, чем у лиц старшего возраста.
Различают упругость на растяжение и упругость на сжатие. Практически для сухожилий наибольшее значение имеет упругость на растяжение, а для су ставных хрящей — на сжатие. По сравнению с упругостью костной ткани упругость сухожилия на растяжение больше в 15 раз, упругость реберного хряща — в 1,5 раза.
Следует заметить, что механические свойства соединительной ткани подвержены большим индивидуальным колебаниям. Они могут быть неодинаковы не только у разных лиц, но и у одного итого же человека, изменяясь в связи с условиями питания и особенностями функционального и возрастного характера.
Мышечные ткани
Основным функциональным свойством мышечной ткани является ее сократимость, которая зависит от способности находящихся в клетках этой ткани сократимых структур изменять свою длину, становясь то короче и толще (сокращение, укорочение), то длиннее и тоньше (расслабление, удлинение).
Существуют три разновидности мышечной ткани: гладкая, поперечнополосатая и сердечная.
Гладкая (неисчерченная) Мышечная ткань. В отличие от других видов мышечной ткани, как показывает изучение микроскопических препаратов, гладкая ткань не имеет поперечной исчерченности. Она находится в стенках кровеносных сосудов, выводных протоков желез, в стенке желудочно-кишечного тракта и многих органов, имеющих полость, а также в толще кожи, образуя Мышцы волос, внутри глазного яблока и др. Структурной единицей этого вида мышечной ткани является миоцит — клетка удлиненной формы, иногда имеющая небольшие отростки. Длина клетки—15—500 мкм, а поперечник —10—20 мкм. Она содержит ядро, располагающееся в центре. Как видно под электронным микроскопом, свойственные поперечнополосатой мышечной ткани сократимые структуры — миофибриллы — здесь отсутствуют. Однако имеются в большом количестве сократительные нити — миофиламенты (протофибриллы) длиной 1—2 мкм и толщиной 5—8 нм (тонкие миофиламенты) и 10—30 нм (толстые миофиламенты). При расслаблении миоцита выявляются только тонкие миофиламенты. Сокращения гладкой мышечной ткани происходят медленно, ритмично и непроизвольно (например, перистальтические движения кишечной трубки, изменение просветов кровеносных сосудов, выводных протоков желез, сокращения мышц радужной оболочки глаза и пр.)
Поперечнополосатая (исчерченная) мышечная ткань. Она названа так благодаря характерно (видимой под микроскопом) исчерченности. Эта ткань участвует в образовании мышц, приводящих в движение Скелет, и поэтому называется еще скелетной мышечной тканью. По функции она является произвольной, поскольку ее сокращения и расслабления подчиняются воле человека. Однако сокращение дыхательных мышц, построенных из этой ткани, может происходить и непроизвольно, как это бывает во время сна. Волокна поперечнополосатой мышечной ткани, являющиеся ее структурными единицами, по своей форме напоминают очень длинные цилиндры, диаметр которых колеблется от 1/100 до 1/10 мм.
Длина волокон составляет от 1 до 40 мм, но может достигать и 10-12см. Каждое мышечное волокно имеет цитоплазму (саркоплазму), двухслойную оболочку (сарколемму) и большое количество ядер вытянутой формы и занимающих обычно поверхностное положение. В саркоплазме находятся митохондрии, от которых зависит энергообеспечение мышечной клетки (рис. 8). Размеры митохондрий 0,3—1,7 мкм х0,2—1 мкм. Они окружены двойной мембраной: наружной, толщина которой 5—10 нм, и внутренней, толщина которой 10—20 нм. Между ними образуется просвет, достигающий около 20 нм. Внутренняя мембрана имеет выросты — кристы, где локализуются ферменты окислительного фосфорилирования.
Саркоплазматическая сеть включает продольные и поперечные трубчатые системы. Продольная система (L-система) имеет вид тонкостенных трубочек, следующих по ходу миофибрилл, анастомозируя друг с другом. Поперечная система (Г-система) представляет собой трубочки (промежуточные пузырьки), расположенные поперечно. Просвет Т-систем связан с внеклеточным пространством, однако продольные и поперечные канальцы не сообщаются друг с другом:

Рис. 8. Схема строения участка поперечнополосатого мышечного волокна по электронномикроскопическим данным
Большой интерес представляет тонкая структура сократительных элементов мышечных волокон — миофибрилл. Каждая миофибрилла проходит через многочисленные тонкие мембраны (телофрагмы или Z-линии), расположенные по отношению к ней в поперечном направлении и представляющие собой продолжение сарколеммы, с которой они непосредственно связаны. Таким образом, создается впечатление, что миофибрилла подразделена на отдельные небольшие сегменты — саркомеры, каждый из которых, в свою очередь, разделен посередине в поперечном направлении чрезвычайно тонкой (более тонкой, чем телофрагмы) перегородкой — мезофрагмой. Как мезофрагмы, так и телофрагмы служат для укрепления миофибрилл, которые через них проходят.
Участки миофибрилл, прилегающие к телофрагме, состоят из светлого, изотропного вещества, а участки, прилегающие к мезофрагме, являются темными, анизотропными.
Таким образом, поперечнополосатая исчерченность обусловлена попеременным чередованием светлых и темных участков миофибрилл, называемых светлыми и темными дисками. С помощью электронного микроскопа установлено, что каждая миофибрилла состоит из протофибрилл, имеющих две разновидности: толстые (16 нм) и тонкие (5—7 нм) нити (см. рис. 8). Толстая нить содержит 180—360 продольно ориентированных молекул белка — миозина. Тонкая нить построена из молекул другого белка — актина, имеющих вид двойной спирали. Между толстыми (миозиновыми) и тонкими (актиновыми) нитями образуются мостики (за счет «головного отдела» молекул миозина).
Сокращение мышечного волокна происходит за счет вхождения нитей актина между нитями миозина (теория скольжения). Саркомер укорачивается, как складная подзорная труба. Объем его остается неизменным, а поперечник увеличивается. Теория скольжения предложена А. Хаксли в 1957 г. Существуют и другие объяснения механизма сокращения мышцы.
Установлена морфологическая и функциональная неоднородность скелетных мышечных волокон. Выделяют волокна первого тип (красные) и волокна второго типа (белые). Красные волокна имеют небольшой диаметр. Они характеризуются высокой активностью окислительных ферментов из-за преобладания аэробных окислительных процессов, высоким содержанием белка-миоглобина. Эти волокна окружены 2—3 кровеносными капиллярами, т. е. уровень кровоснабжения у них высокий. Красные мышечные волокна относят к медленным, тоническим.
Белые волокна более толстые. Они содержат в больших количествах фосфорилазу и АТФ, обеспечивающие анаэробные процессы. Основной источник энергии — Гликоген. Уровень кровоснабжения у них ниже, чему красных волокон: на одно мышечное волокно приходится в среднем один кровеносный капилляр. Белые волокна считаются быстрыми, тетаническими.
Существует также переходный тип мышечного волокна.
Электронно-микроскопически и гистохимические волокна указанных типов различаются по содержанию митохондрий, липидов и соотношению окислительных ферментов с ферментами гликолиза, а также по ширине Z-линий между саркомерами. В красных волокнах (медленных) много митохондрий и липидов (они — основной источник энергообеспечения); окислительные ферменты преобладают над ферментами гликолиза; Z-линия широкая. В белых волокнах (быстрых) мало митохондрий, липидов и окислительных ферментов, но много гликолитических ферментов; Z-линия узкая. Существенных отличий в содержании гликогена и строении миофибрилл в волокнах этих типов не установлено.
Сравнение ряда мышц конечностей выявило мозаичный характер в распределении волокон первого и второго типов.
Оболочка волокна — сарколемма — способствует укреплению взаимосвязи мышечных волокон. Внутренний ее слой — плазмо лемма — аналогичен оболочкам других клеток. Наружный слой — базальная мембрана — состоит из тонких фибрилл и тесно связан с окружающей волокно соединительной тканью — эндомизием. Строение мышцы как органа см. на стр. 127.
Поперечнополосатые мышечные волокна вместе с иннервирующим их нейроном составляют двигательную единицу. В ее состав входят красные или белые мышечные волокна. Их число весьма значительно (например, в медиальной головке икроножной мышцы — 1634, а в передней большеберцовой мышце — 667).
В месте подхода аксона двигательной нервной клетки к мышечному волокну образуется нервно-мышечный синапс в виде двигательной бляшки. Окончания аксона лежат в углублениях на поверхности мышечного волокна, выстланных сарколеммой. Аксон нервной клетки не проникает внутрь мышечного волокна. Между ними остаются синаптические щели шириной 20—60 нм. Здесь импульсы, побуждающие мышцу к сокращению, передаются от аксона нервной клетки на мышечное волокно (см. рис. 9). При этом из саркоплазматической сети последнего Ионы кальция поступают в миофибриллы. В результате Актин приобретает способность взаимодействовать с миозином, что приводит к сокращению волокна (см. стр. 36).
Сердечная мышечная ткань. Эта ткань является поперечнополосатой, но имеет особенности строения, позволяющие выделить ее в отдельную группу. Одна из этих особенностей —то, что клетки сердечной мышечной ткани, кардиомиоциты, образуют между собой многочисленные соединения. Сокращения сердечной мышцы происходят непроизвольно.
Ядра кардиомиоцитов занимают центральное положение, миофибриллы располагаются по периферии клетки. В отличие от скелетной мышечной ткани в клетках сердечной мышечной ткани намного больше митохондрий, с большим количеством крист. Это свидетельствует о хорошем энергообеспечении сердечной мышцы. Саркоплазматическая сеть здесь, наоборот, хуже развита, чем в скелетной мышечной ткани.
Контакты клеток сердечной мышцы происходят в области вставочных дисков и боковых выростов цитоплазмы. Вставочные диски не только соединяют клетки между собой, но и участвуют в передаче возбуждения от одной клетки к другой. Боковые выросты и вставочные диски обеспечивают сокращение миокарда как единого целого. Среди клеток, обеспечивающих сокращение, есть такие, которые проводят импульсы внутри сердца. Это большие клетки, богатые саркоплазмой, с малым количеством миофибрилл и митохондрий, с крупным ядром, не всегда расположенным в центральной части клетки.
Нервная ткань состоит из нервных клеток с их отростками и окончаниями этих отростков. К ней относятся также образования, имеющие для собственно нервной ткани опорное и трофическое значение и именуемые нейроглией /макро- и микроглия/.
Каждая нервная клетка содержит цитоплазму и ядро округлой или слегка овальной формы. В цитоплазме хорошо выражены митохондрии и комплекс Гольджи. Встречаются отложения тигроида, возникшие на основе видоизменений эндоплазматической сети. Под электронным микроскопом определяются нейрофиламенты — нити толщиной около 10 нм.
Нервная клетка с ее отростками называется нейроном (рис. 9). Он представляет собой структурную единицу нервной ткани. Нервные клетки, отростки которых идут к органам (например, мышцам) и несуткним импульсы, побуждающие их к деятельности, называются двигательными, выносящими, эфферентными. Нервные клетки, отростки которых проводят импульсы от периферии к центру, являются чувствительными, приносящими, афферентными. Кроме двигательных и чувствительных нейронов есть огромное количество вставочных нервных клеток, связывающих друг с другом чувствительные и двигательные Нейроны. Величина тел нервных клеток разнообразна и составляет в поперечнике от 25 до 150 мкм. Существуют нервные клетки с телами настолько больших размеров (например, клетки передних рогов спинного мозга), что они находятся на грани видимости невооруженным глазом. Форма тел нервных клеток тоже разнообразна: многоугольная, веретенообразная, продолговатая, круглая. Она связана с количеством отростков клетки, так как соответственно каждому отростку тело нейрона образует выступ.

Рис. 9. Строение двигательного нейрона:
1 — перикарион; 2 — аксон и нервное волокно; 3 — нервные окончания в мышце; 4 — дендриты; 5 — мякотная оболочка; 6 — перехваты Ранвье. В схеме сопоставлены световая и Электронная Микроскопия (по Г.Ф. Иванову и Ковальскому, изменено)
Характер отхождения отростков позволяет дифференцировать нейроны на униполярные (с одним отростком, Т-образно делящимся на две ветки), биполярные (с двумя отростками) и мультиполярные (многоотростчатые).
Одни отростки нервных клеток — короткие, протоплазматические, древовидно разветвляющиеся — дендриты; другие — длинные нейриты, или аксоны. Длина отростков нервных клеток может быть очень велика (в некоторых местах более 1 м). По нейритам раздражения идут от тела клетки, в то время как по дендритам — к телу клетки.
Отростки продолжаются в составе нервных волокон в виде осевых цилиндров, обычно покрытых глиальными оболочками более простого или сложного строения. Только в сером веществе головного мозга у отростков нервных клеток оболочек нет. Отростки нервных клеток, составляющие Белое вещество центральной нервной системы, окружены оболочкой.
Отростки нервных клеток образуют нервные окончания: рецепторы (у дендритов) и эффекторы (у нейритов). Нервные клетки соединяются друг с другом и с иннервируемым органом (мышцей, кожей или железой) при помощи особых аппаратов, именуемых синапсами, имеющими крайне разнообразные формы.
Отростки клеток служат не только для проведения нервных импульсов, но и для транспортировки белков и других веществ от тела или к телу нейрона. Существуют два тока внутриклеточных продуктов: медленный со скоростью 1—2 мм в сутки и быстрый — 5—10 мм в час.
Известно, что некоторые нейроны обладают способностью к нейросекреции. Их называют секреторными. Образование секрета связано с тигроидной субстанцией и комплексом Гольджи. Гранулы нейросекрета перемещаются по аксону от тела клетки, однако поступают не в область синапсов, как остальные транспортируемые вещества, а в кровь или в спинномозговую жидкость (подобно гормонам).
Нервные волокна варьируют по своему диаметру. Одна часть осевых цилиндров располагается в «футляре» из миелина (жироподобного вещества) — это миелиновые волокна, или мякотные; другая часть лишена такого покрова — это амиелиновые, или безмякотные, волокна. Миелиновая оболочка увеличивает скорость проведения нервных импульсов, следовательно, ее наличие функционально оправдано. Безмякотные волокна образуют полиаксональную оболочечную систему, аксоны которой окружены клетками-сателлитами (шванновскими). При образовании миелиновых волокон аксон вначале располагается на периферии клетки-сателлита, затем «вдавливается» в нее, что приводит к образованию «брыжейки» — мезаксона (рис. 10). Мезаксон спирально разрастается вокруг аксона, и в местах соприкосновения складок разросшегося мезаксона образуется миелин. По ходу мякотного волокна миелиновый покров местами истончается, образуя перехваты Ранвье. Это биологически активные участки нерва, где скапливаются митохондрии, многие ионы и продукты метаболизма.
Нервные волокна, покрытые слоем соединительной ткани — эндоневрием, образуют пучки, окруженные более толстыми прослойками периневрия, и формируют нерв. В зависимости от количества волокон и пучков, входящих в состав нерва, толщина его сильно варьирует—от тонких ветвей, невидимых невооруженным глазом, до очень толстых (например, толщина седалищного нерва может быть более 1 см).
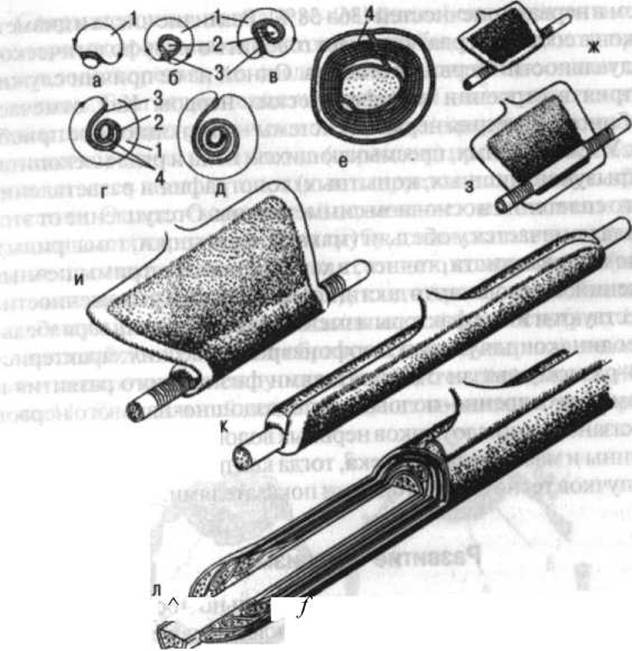
Рис. 10. Схема миелинизации нервного волокна: а—е — последовательные стадии вращения мезаксона вокруг аксона на поперечном срезе; ж—л — вид процесса; 1 — шванновская клетка; 2 — аксон; 3 — мезаксон; 4 — слои миелина (по Герен)
Существуют два крайних варианта строения периферического нерва: малопучковый (нерв тонкий, состоит из небольшого количества крупных пучков при компактном расположении волокон в пучке) и многопучковый (нерв толстый, образующие его пучки небольшого диаметра, расположение волокон в пучке рыхлое). Количество волокон в составе отдельных нервов весьма изменчиво: срединный нерв на уровне середины плеча содержит 19—32 тыс. волокон, локтевой на том же уровне —13—18 тыс., мышечно-кожный — 3—12 тыс. Нервные стволы отличаются по диаметру слагающих их волокон: мелкие и средние миелиновые волокна составляют в срединном нерве 11-45%, в локтевом - 9-37%, в лучевом - 10-27%. В нервах, иннервирующих кожу, этих волокон больше (60—80%), чем в иннервирующих мышцы (18—40%); в межреберных нервах больше (70—80%), чем в нервах конечностей (36—38%). Различие числа и диаметра волокон в составе нерва позволяет говорить о «морфологической индивидуальности» нервных стволов. Одной из ее причин служит Асимметрия в строении периферических нервов. Как отмечает В.В. Бобин, асимметрия нервной системы — эволюционное приобретение. У земноводных, пресмыкающихся, птиц и ряда млекопитающих (грызунов, хищных, копытных) топография и разветвление плечевого сплетения в основном симметричны. Отступление от этого правила намечается у обезьян (макаки, мартышки, гамадрилы), особенно в нервах кисти, точнее, в характере их внутримышечных разветвлений. У человека это достигает наибольшей выраженности.
Существуют и иные факторы изменчивости. Уровень вариабельности неодинаков для разных морфоневрологических характеристик и по-разному связан с показателями физического развития и возрастом. Для бедренно-полового и подвздошно-пахового нервов было показано, что число пучков нервных волокон не зависит от возраста, длины и массы тела человека, тогда как площадь поперечного сечения пучков тесно связана с этими показателями.
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.