БІОФІЗИКА РОСЛИН - Ю. І. Посудін - 2004
IІ. ПРОЦЕСИ ПЕРЕНОСУ В СИСТЕМІ РОСЛИНА-ҐРУНТ-ПОВІТРЯ
12. ВПЛИВ ЗОВНІШНІХ ФАКТОРІВ НА РОСЛИНИ
12.2. ФОТОБЮЛОТІЧНІ РЕАКЦІЇ РОСЛИН
12.2.1. Класифікація фотобіологічних реакцій рослин
Рослині для розвитку потрібні сонячне світло, двоокис вуглецю, Вода та поживні речовини, завдяки яким відбуваються розвиток рослини - якісні зміни в структурі і функціональній активності рослини та її частин (клітин, тканин та органів) у процесі життєвого циклу рослини, а також Diem рослини - незворотне збільшення розмірів рослин або її органів, зумовлене новоутворенням клітин, органів або їх окремих елементів. Для багатьох процесів росту та розвитку рослини найважливішим зовнішнім фактором є світло. Під впливом сонячного випромінювання відбуваються фотобіологічні реакції - фізичні або хімічні зміни в рослинних системах. Усі фотобіологічні реакції можна поділити на чотири основні групи [Конев и Волотовский, 1979]:
1. Енергетичні - реакції, в ході яких світлова енергія внаслідок синтезу нових органічних молекул трансформується в хімічну. Прикладом цих реакцій може бути Фотосинтез.
2. Інформаційні - реакції, при яких світло є керуючим сигналом, що викликає через спеціалізовані механізми утворення фотопродуктів та забезпечує інформацію щодо навколишнього середовища. До інформаційних можна віднести фоторух, фотоморфогенез, фототропізм, Фотоперіодизм.
3. Біосинтетичні - реакції, при яких у складному ланцюзі послідовних етапів синтезу органічних молекул є окремі фотохімічні стадії, що протікають лише під впливом світла. До цих реакцій відносяться біосинтез хлорофілу, індукція синтезу пігментів та вітамінів.
4. Деструктивно-модифікуючі реакції, пов'язані з пошкодженням світлом молекул біологічного субстрату, що призводить до летальних або мутаційних наслідків. До цього типу реакцій відносяться фотосенсибілізація, фотореактивація, відгук біологічних систем на ультрафіолетове випромінювання.
Розглянемо деякі важливі для вищих та нижчих рослин фотобіологічні реакції.
12.2.2. Фотосинтез
Основними функціональними одиницями, що забезпечують фотосинтетичну активність рослин і водоростей, є фотосистеми ФСІ і ФСІІ, які містять близько 250-400 молекул пігментів. Ці пігменти здатні поглинати кванти світла, але лише одна молекула хлорофілу даної фотосистеми спроможна перетворювати поглинуту енергію для фотохімічних реакцій. Ця молекула хлорофілу називається реакційним центром фотосистеми, тоді як інші - антенами. Після поглинання кванта світла антенними пігментами поглинута енергія переноситься до реакційних центрів фотосистем, що супроводжується збудженням молекули хлорофілу Р і переходом електронів на більш високий енергетичний рівень. Молекула Р*680 у збудженому стані переносить електрони акцептору - феофітину Фф, після чого - первинному хіноновому акцептору QA і вторинному хіноновому акцептору QB. Далі електрони послідовно передаються на пул молекул пластохінона ПХ, що переносить через ліпідну фазу мембрани електрони і протони, залізо- сірчаний білок FeSR, цитохром/і пластоціанін Р. Електрони переходять по електронно-транспортному ланцюгу до ФСІ, де енергія світла передає електрони від молекули хлорофіла Р700 до електронного акцептора Р430 після чого відбувається подальше перенесення електронів через фередоксин Fd до кофермента НАДФ (рис. 12.5).
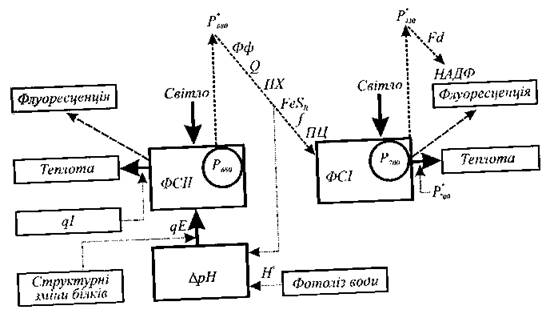
Рис. 12.5. Схема, що ілюструє процеси перетворення енергії у флуоресценцію і тепло під час нециклічного транспортування електронів.
Крім того, поглинута енергія може бути звільнена у вигляді теплоти або витрачена на такий випромінювальний процес, як флуоресценція. При звичайних температурах флуоресценція хлорофілу походить від пігментів антени ФСІІ; вклад ФСІ у флуоресценцію хлорофілу має місце лише в довгохвильовій частині спектра (740 нм). Змінна флуоресценція в основному походить від ФСІІ, а перенесення збудження до ФСІ можна трактувати як додатковий конкуруючий шлях дезбудження ФСІІ. Під час функціонування фотосинтетичного апарата випромінювання флуоресценції хлорофілу а становить 2-5 % поглинутої енергії і залежить від типу хлоропласта, віку, фізіологічного стану рослини, стресових умов, інтенсивності та довжини хвилі збуджуючого випромінювання.
Перехід електронів уздовж електронно-транспортного ланцюга супроводжується зменшенням (гасінням) флуоресценції хлорофілу. Це зменшення відбувається завдяки окисленню акцептора, який являє собою комплекс феофітину і хінонів. Якщо акцептор окислюється завдяки перенесенню електронів до НАДФ і, врешті-решт, до СО2 флуоресценція зменшується. Такий процес називається "фотохімічним гасінням". Характеризується цей процес коефіцієнтом фотохімічного гасіння qP. У той же час існують інші механізми гасіння нехімічної природи або "нефотохімічного гасіння", які характеризуються коефіцієнтом нефотохімічного гасіння qN. Основними з процесів гасіння є залежне від енергії гасіння, пов'язане з індукованим протонним градієнтом крізь тилакоїдну мембрану, та гасіння, пов'язане з фотоінгібуванням, що викликається надлишковим опромінюванням. Отже, флуоресценція є комплементарним процесом по відношенню до фотохімічних та теплових процесів: вихід флуоресценції тим більший, чим менші розтрати енергії на фотохімічні реакції або теплоту.
12.2.3. Фоторух водоростей
Рух або зміна руху організмів, що викликаються світлом, називається фоторухом. Фоторух є результатом фоторегуляцїї руху - сукупності елементарних процесів, індукованих світловим стимулом, а саме: фоторецепції, сенсорного перетворення світлового стимулу в фізіологічний сигнал, який керує роботою рухового апарата, що реалізує фотоорієнтацію організму. Світло - це важливий зовнішній фактор, який завдяки фоторуху використовується рухливими організмами (в першу чергу - водоростями) для пошуків оптимальних умов існування.
Фоторецепція. Розглянемо основні механізми фоторецепції водоростей [Посудин, 1989; Масюк и Посудин, 1991].
Періодичне освітлення та затінювання стигмою фоторецептора. В процесі обертання клітини навколо своєї поздовжньої осі виникає амплітудна модуляція світла, що потрапляє на фоторецептор. Величина модуляції залежить від напрямку руху клітини відносно напрямку поширення стимулюючого світла. Роль модулятора виконує специфічна органела - стигма. Модуляційний механізм притаманний представникам Euglenophyceae.
Дихроїчний механізм. У його основі лежить анізотропія поглинання світла дипольними фоторецепторними молекулами, що орієнтовані певним чином відносно поздовжню! осі клітини. Максимальне поглинання світла відбувається під час руху клітини паралельно напрямку поширення світла. Дію такого механізму знайдено у евгленофітової водорості Euglena gracilis, для якої не виключається поєднання модуляційного та дихроїчного механізмів.
Хвильоводний механізм. Передбачається поширення бокового світла, перпендикулярного поздовжній осі клітини, через спарені тилакоїди, розташовані перпендикулярно до площини пігментованого шару стигми та поздовжньої осі клітини. Ці тилакоїди, що мають високий показник заломлення відносно проміжків між ними, утворюють специфічний світловід для світла, що поширюється. Такий механізм знайдений у криптофітових водоростей Chroomonas Hansgh та Cryptomonas Ehr.
Оцелоїдний механізм. Фоторецепція відбувається завдяки спеціалізованій органелі - оцелоїду, який являє собою своєрідну фокусуючу систему. Світло, що потрапляє на оцелоїд, фокусується кришталевим тілом на ламелярному шарі ретиноїдного тіла. Подібний механізм притаманний представникам динофітових водоростей із родини Wamowiaceae.
Інтерференційний механізм. Стигма, яка складається з кількох пігментованих та непігментованих шарів та, можливо, розташована під фоторецептором, виконує функції багатошарової чвертьхвильової пластини. Внаслідок відбивання світла від кожного шару має місце Інтерференція світла; якщо товщина шарів та проміжків між ними дорівнює чверті довжини світлової хвилі, відбувається підсилення світла, що потрапляє на фоторецептор. Такий механізм спостерігається у представників Chlorophyceae, Prasinophyceae та у деяких видів Dinophyceae.
Дифракційний механізм. У багатьох зелених водоростей стигма містить лише один шар пігментованих глобул. Передбачається, що пігментовані глобули стигми являють собою періодичну структуру, яка виконує функції дифракційної решітки. Під час взаємодії світла з такою структурою відбувається дифракція світла, що супроводжується утворенням дифракційних максимумів. Інтенсивність та просторове положення цих максимумів залежать від геометрії періодичної структури, кута падіння світла на неї та довжини світлової хвилі. Залежно від збігу дифракційного максимуму з місцезнаходженням рецептора змінюється величина світлового сигналу, що потрапляє на фоторецептор.
Будова деяких фоторецепторних систем водоростей пояснюється на рис. 12.6.
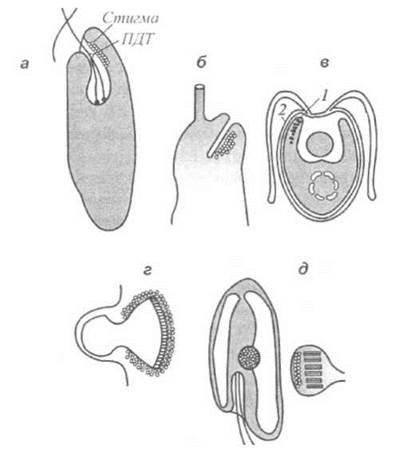
Рис. 12.6. Будова деяких фоторецепторних систем водоростей: а - стигма та параджгутикове тіло евгленофітової водорості Euglena gracilis; б - параджгутиковий виступ у жовто-зелених водоростей в - стигма (7) та здогадний рецептор (2) зелених водоростей; г - рецептор криптофітових водоростей; д - оцелоїд (родина Warnowiaceae).
Сенсорне перетворення. Після поглинання фоторецептором кванта світла має місце низка подій, яка призводить до перетворення світлового стимулу в фізіологічний сигнал, який керує роботою рухового апарата водорості - так званого сенсорного перетворення. Є багато методологічних та експериментальних прийомів, що ставлять своєю метою освітлення основних етапів сенсорного перетворення - це застосування специфічних метаболічних препаратів (іонів хімічних елементів, блокаторів кальцієвих каналів, іонофорів, іонотропних препаратів тощо). На жаль, до цього часу проблема з'ясування проходження світлового сигналу від фоторецептора до рухового апарата являє собою своєрідну "чорну скриньку", оскільки багато аспектів цієї проблеми поки що не розв'язано.
12.2.4. Фоторух хлоропластів
Індукований світлом рух хлоропластів у клітині здійснюється так: по-перше, хлоропласти можуть брати участь у загальному русі цитоплазми (поступальному, обертальному, циркуляційному); при цьому світло, впливаючи на частину цитоплазми, викликає зміну швидкості її руху, тоді як просторове взаємне розташування хлоропластів залишається незмінним. Така світлозалежна реакція хлоропластів називається фотодінезом. По-друге, хлоропласти здатні переорієнтовуватися в межах клітини; результуючий розподіл їх пов'язаний з характером поглинання світла клітиною, яке визначається напрямком поширення світла. Така реакція отримала назву фотоорієнтації хлоропластів. Якщо в першому випадку стимулом є величина інтенсивності світла, то в другому - напрямок поширення світла.
Фотодінез демонструють хлоропласти Elodea та Vallisneria. Зміна швидкості потоку цитоплазми в епідермальних клітинах Elodea та мезофільних і епідермальних клітинах Vallisneria під впливом світла викликає переміщення хлоропластів. Існує гіпотеза, згідно з якою світло керує активацією актиноподібних мікрофіламентів та зміною рівня АТФ. Індуковане синім світлом збільшення окислювального фосфорилування завдяки флавінам вважається відповідальним за зміну пасивної рухливості цитоплазми при низькій швидкості потоків. При середній швидкості починається конкуренція з боку фотосинтезу; при великій швидкості, коли фотосинтез досягає насичення, має місце надлишок АТФ за рахунок циклічного фосфорилування, який разом з індукованим світлом окисленням АТФ призводить до активації актоміозину. На користь цієї гіпотези свідчать експерименти з застосуванням роз'єднувачів або інгібіторів фотосинтетичного електронного потоку.
Щодо орієнтації хлоропластів, то тут слід виділити такі варіанти: під впливом бокового низькоінтенсивного освітлення хлоропласти переміщуються до стінок клітини і розташовуються перпендикулярно до напрямку поширення світла (епістрофізм); при збільшенні інтенсивності світла хлоропласти орієнтуються уздовж бокових стінок клітин за напрямком поширення світла (парастрофізм). У темряві хлоропласти орієнтуються хаотично (апострофізм). Схематично ці типи орієнтації хлоропластів наведено на рис. 12.7. На характер орієнтації хлоропластів впливають розсіювання та поглинання світла усередині клітини, що викликають появу світлових градієнтів; "лінзові" ефекти, які призводять до фокусування світла. Одним з основних (але далеко не простих) аспектів проблеми фоторуху хлоропластів є визначення структури, пігментного складу та місцезнаходження фоторецептора. Деякі організми (Lemna, Vallisneria, Vaucheria, Mesotaenium, Selaginella, Mougeotia, Hormidium, Spirodela) демонструють реакцію хлоропластів на синє світло, рецепторами якого можуть бути флавопротеїди. Рух хлоропластів при опромінюванні інтенсивним червоним світлом можна спостерігати у Selaginella та Vallisneria, а під впливом низькоінтенсивного червоного світла - у Hormidium. Здійснюють рецепцію червоного світла Фотосинтетичні пігменти, про що свідчать експерименти з препаратами, які впливають на фотосинтез. Реакцію на червоне світло демонструють хлоропласти Mesotaenium та Mougeotia; фоторецепторним пігментом у даному випадку є фітохром. Слід відзначити також і різноманіття розташування фоторецепторів - вони можуть знаходитися не лише в хлоропластах, а також у цитоплазмі або плазматичній мембрані.
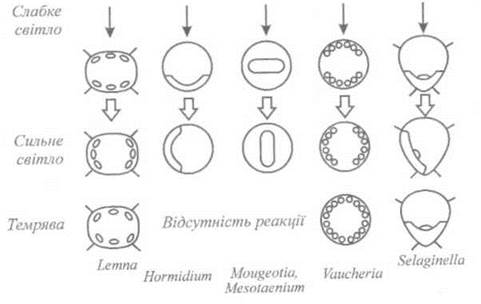
Pис. 12.7. Типи фотоорієнтації хлоропластів.
12.2.5. Фотоморфогенез
До цього типу фотобіологічних реакцій можна віднести всю сукупність контрольованих світлом процесів, пов'язаних з розвитком та ростом рослини, причому реакції рослини не характеризуються орієнтованою відносно світла направленістю, а світловий стимул не має просторової направленості та періодичності.
Фотоморфогенетичними реакціями можна вважати проростання насіння, видовження стебла, формування листя, розвиток хлоропластів тощо.
Більшість фотоморфогенетичних реакцій рослин відбувається завдяки участі трьох типів фоторецепторів - фітохрому, криптохрому - рецептора синього та ультрафіолетового УФ-А (320-400 нм) випромінювання. Фітохром складається з білкової частини та хромофора - лінійного тетрапіролу (фітохромобіліну), що поглинає світло. Фітохром може існувати у двох різних формах, які можуть перетворюватися одна в одну: Рч (фізіологічно неактивна форма, що поглинає червоне світло) та Рдч (фізіологічно активна форма, що поглинає дальнє червоне світло). Коли молекула Рчпоглинає фотон червоного світла з довжиною 660 нм, вона перетворюється у Рдч; якщо молекула Рдч поглинає фотон дальнього червоного світла з довжиною 730 нм, вона перетворюється у Рч. Такі переходи під впливом світла називаються фотоконверсіями. Участь фітохрому в морфогенетичних процесах підтверджують досліди по оборотній стимуляції та інгібуванню морфогенетичних процесів червоним (660 нм) та дальнім червоним (730 нм) світлом. Фітохром локалізований у мембранах рослини; основним механізмом дії фітохрому слід вважати регуляцію функцій мембран за рахунок конформаційних змін фітохрому, що впливають на проникність мембрани, з якою він зв'язаний, на активний транспорт іонів через мембрану та активність фітогормонів і білків, зв'язаних з мембраною. Криптохром відповідає за поглинання світла в синій (400-500 нм) та ультрафіолетовій УФ-А областях спектра; функції його можуть виконувати флавіни або каротиноїди. Складається криптохром з білка, зв'язаного з двома хромофорами. Спектр дії криптохрому займає область 390-480 нм з максимумом при 450 нм. Криптохром відповідає за інгібування видовження стебла, розвиток листя, фотоперіодичне цвітіння тощо.
12.2.6.Фототропізм
Індукований світлом рух частини фіксованого рослинного організму називається фототропізмом. Ця фотобіологічна реакція являє собою відгук рослини на градієнт світла. Такі градієнти можуть утворюватися за рахунок поглинання або розсіювання світла, яке проходить через рослинний орган. Розрізняють позитивний (рух до джерела світла) та негативний (від джерела) фототропізм. Для верхівок паростків характерним є позитивний фототропізм, тоді як для коріння - негативний. Якщо, наприклад, колеоптиль вівса освітити світлом низької інтенсивності з одного боку, або світлом різної інтенсивності з різних боків, то цей орган вигинається у сторону більш яскравого світла. Типовий спектр дії фототропізму (рис. 12.8) характеризується максимумами у видимій області спектра при 420 нм, 450 нм та 475 нм; крім того, є максимум при 370 нм в ультрафіолетовій області спектра. Таким чином, спектр дії фототропізму свідчить про те, що лише синє світло та частина ультрафіолетового випромінювання викликають утворення вигинів.
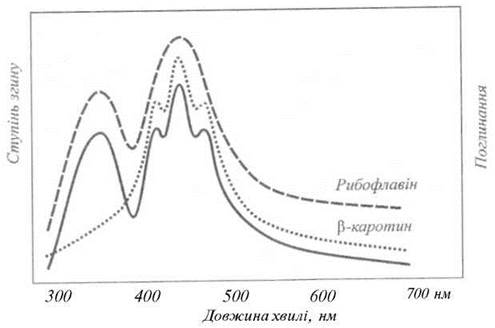
Рис. 12.8. Типовий спектр дії фототропізму.
Імовірними хромофорами, що відповідають за фототропні реакції, є каротиноїди та флавіни (або флавопротеїни), про що свідчить збіг спектра дії фототропізму зі спектрами поглинання цих пігментів.
Слід зауважити, що питання щодо природи хромофорів ще далеко від остаточного розв'язання. Фототропні вигини рослинного органа пов'язані з асиметричним розподілом у цьому органі ауксину - фізіологічно активної речовини, яка утворюється в клітинах рослини і регулює процеси росту. Світло викликає вигин органа, впливаючи на латеральний розподіл ауксину. Різниця в концентрації ауксину обумовлює, в свою чергу, неоднакову швидкість росту органа з двох сторін, що й призводить до вигину.
Механізм дії ауксину пов'язаний з двома факторами [Гапстон и др., 1983] - розтяжністю клітинних оболонок та тургорним тиском клітинної рідини, що діє на цю оболонку. Ауксин викликає збільшення поздовжніх розмірів клітин, що обумовлено спіральним розташуванням целюлозних мікрофібрил у клітинній оболонці та тим, що ауксин збільшує розтяжність цієї оболонки. Таке індуковане ауксином збільшення пластичної розтяжності клітинної оболонки спостерігається лише в живих клітинах. Є гіпотеза, що для забезпечення відносного сковзання целюлозних ланцюгів необхідно розірвати поперечні зв'язки між молекулами целюлози; ці функції виконує поки ще невідомий фермент, активність якого залежить від рівня pH внутрішньоклітинного середовища. Коли під впливом ауксину, присутнього в клітині, іони водню накачуються в клітинну оболонку, фермент активізується і руйнує поперечні зв'язки; целюлозні ланцюги звільняються та отримують можливість ковзати один відносно одного. Таке ковзання відбувається під впливом тургорного тиску клітинної рідини, що призводить до розтягування оболонки та збільшення розмірів клітини.
12.2.7. Фотоперіодизм
Ритмічні зміни різноманітних морфологічних, біохімічних та фізіологічних властивостей і функцій організмів під впливом чергування та тривалості світлових і темнових інтервалів називаються фотоперіодизмом.
Згідно з фотоперіодичними реакціями рослини можна поділити на три групи: рослини короткого дня (зацвітання та плодоношення настають при скороченні дня до 8-12 год.); рослини довгого дня (той же ефект досягається подовженням дня до 16-20 год.) та нейтральні рослини (в яких зміна дня не спричиняє помітних змін протягом життєвого циклу). До групи короткого дня відносяться бавовник, кофе, полуниця, просо, рис, соя, тютюн, цукрова тростина; до групи довгого дня - жито, льон, люпин, овес, пшениця, редиска, цукровий буряк, шпинат, ячмінь; представниками нейтральних рослин є горох, гречка, соняшник.
Від фотоперіоду залежать такі процеси, як цвітіння, спокій бруньок, скидання листя, проростання насіння, формування цибулин та бульб тощо. Характер фотоперіодичної реакції пов'язаний з географічним походженням рослини і сформувався протягом еволюції. Відсутність точних даних щодо спектрів дії фотоперіодизму не дозволяє сказати щось певне відносно природи фоторецептора, який відповідає за фотоперіодичну реакцію. Серед існуючих гіпотез [Гродзинский, 1972; Конев и Волотовский, 1979] слід відзначити фітохромну (яка пов'язує фотобіологічну реакцію з фотоперетворюваннями фітохрому), гормональну (фотобіологічний ефект викликається утворенням особливого гормону флоригену), гіпотезу ендогенних ритмів (що базується на взаємодії біологічних та астрономічних годинників), інгібіторну гіпотезу (що припускає утворення в рослині при несприятливих фотоперіодичних умовах особливих інгібіторов цвітіння). Жодна з цих гіпотез поки що не спроможна пояснити всю складність та різноманіття фотоперіодичних реакцій рослин.
12.2.8. Фотосенсибілізація
Явище фотосенсибілізації. У 1900 р. Оскар Рааб вивчав вплив дії світла та акридинових барвників на життєдіяльність найпростіших (Paramecium). Він помітив, що ні світло окремо, ні барвник не впливали на інфузорії, тоді як сумісна дія цих двох факторів призводила до їхньої загибелі. Явище фотосенсибілізації - це процеси, при яких світлова енергія, поглинута молекулами, що мають хромофори (сенсибілізатори), передається іншим молекулам, неспроможним самостійно поглинати світло.
Це явище поширене у природі - відомо, що деякі дикорослі рослини містять сильнодіючі хімічні сполуки; тварини, які поїдають ці рослини, стають чутливими до світла в непігментованих ділянках шкіри, особливо навколо очей, рота, вух та копит, де волосяний покрив рідкий. Основні симптоми - свербіж, виникнення пухирців розміром з горошину (які лопаються), Кон'юнктивіт, стоматит, гарячковий стан, запальний процес (навіть у мозку), збудження або пригнічення. Тварина починає тертися, вносить інфекцію і може загинути через 8-10 год.
Основні механізми фотосенсибілізації. Можна виділити процеси, які характеризують взаємодію світла, сенсибілізатора та біологічного субстрату, а саме: поглинання світла молекулою сенсибілізатора та перехід її у збуджений стан; інтерконверсія у триплетний стан; перенос енергії триплетного стану сенсибілізатора до триплетного стану кисню; перенос енергії триплетного стану сенсибілізатора до молекули субстрату з утворенням радикалів. Реактивні властивості синглетного кисню і хімічних радикалів спричиняють певні зміни молекул субстрату.
Основні типи фотосенсибілізацій. Всі захворювання, викликані фотосенсибілізацією, можна поділити натри класи.
Первинна фоточутливість. У ролі хімічних сполук, які підвищують чутливість тварин до світла (фотосенсибілізаторів), виступають гіперицин та фагопірин. Гіперицин - червоний пігмент, що міститься у листках, стеблинах та квітках представників родини Guttiferae, зокрема: Hypericum perforatum (звіробій), H.crispum, H.pulchrum, H.leucopty codes, H.maculatum. Загальна назва захворювань - гіперицизм. Тварини, які поїдають рослини, що містять гіперицин, стають чутливими до світла. Зазвичай під вплив гіперицину потрапляють вівці, коні, велика рогата худоба Хоча кози й свині також можуть потрапити під фотосенсибілізуючу дію гіперицину. У овець, які поїдають Hypericum perforatum, симптоми не проявляються, коли вони знаходяться у темному приміщенні. Але на світлі тварини проявляють фоточутливість, що триває кілька тижнів. Цікавим є факт, що багато комах не наближаються до рослин, які містять гіперицини. Винятком є бджола Chrysolina gemellata, що живиться нектаром Hypericum perforatum. Але вона не відвідує інші рослини, контролюючи таким чином утворення гіперицину на певних ареалах.
Фагопіризм викликається спорідненим гіперицину рослинним пігментом фагопірином, який присутній у представників Fagopyrum (F.esculentum або Poligonumfagopyrum) - гречки посівної. Фагопірин також має фотосенсибілізуючий вплив на свиней, худобу, курчат. Можливі захворювання овець, свиней, коней (які поїдають на сонці гречку, просо, шведську конюшину) на шкірну екзантему, яка відсутня на пігментованих ділянках тіла; у овець, коней і великої рогатої худоби з білими мітками виникають дерматити, стоматити, а також Опіки на місцях, позбавлених пігментації.
Фоточутливість, пов'язана з порушенням синтезу пігментів. Серед захворювань цього типу слід виділити природжену порфірію, що викликається ендогенними порфіринами - уропорфірином та копропорфірином. Перший виділяється у сечу, а другий відкладається в кістках та зубах (звідки походить ще одна назва захворювання - рожеві Зуби). Є гіпотеза, згідно з якою ці порфірини виникають внаслідок порушення синтезу протопорфірину, який є частою молекули гемоглобіну. Але в цілому походження порфірії невідоме.
Гепатогенна фоточутливість. Слід звернути увагу на фотосенсибілізуючі властивості філоеритрину - порфірину, одержаного з хлорофілу за допомогою бактерій, що є в кишечнику різних тварин, за рахунок вилучення фітольної групи, що перешкоджає проходженню хлорофілу через клітинні мембрани. У нормальному стані Організм травоїдних позбавляється від філоеритрину за допомогою жовчі; коли ж тварина поїдає деякі трави, що блокують жовчний протік, філоеритрин потрапляє у Кров і шкірний покрив. У цьому разі він починає діяти як сенсибілізатор при опромінюванні тварини видимим або ультрафіолетовим світлом. Шкірний покрив набуває жовтого забарвлення, набрякає. Філоеритрин може бути виділений із крові хворих тварин і кристалізований. При ін'єкції його тваринам, які поїдали зелений корм, філоеритрин викликає симптоми жовтяниці. При з'їданні деяких зелених кормів тварина стає чутливою до світлового впливу.
Серед рослин, які викликають захворювання тварин, можна відзначити представників роду Tribolus ( звідси назва хвороби - триболізм) зокрема Т.terrestris та Т.ubis (родина Zygophyllaceae); деякі види Lippia: L.rhenanni, L.pretoriensis (родина Verbenaceae); трави Pannicum; P.laevifolium та P.coloratum (родина Graeminae). У наших широтах фотосенсибілізацію викликають конюшина (Trifolium L.) та сухоребрик високий (Sysimbrium altissimum L.).
Запобігання хвороб - зміна пасовищ, утримання худоби в затемнених місцях.
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.