БІОФІЗИКА РОСЛИН - Ю. І. Посудін - 2004
IІ. ПРОЦЕСИ ПЕРЕНОСУ В СИСТЕМІ РОСЛИНА-ҐРУНТ-ПОВІТРЯ
12. ВПЛИВ ЗОВНІШНІХ ФАКТОРІВ НА РОСЛИНИ
12.1. ВПЛИВ ГРАВІТАЦІЇ НА РОСЛИНИ
12.1.1. Гравітаксис водоростей
Вертикальна міграція водоростей являє собою поведінковий механізм пошуків оптимального з точки зору їхнього росту та репродукції положення у водному середовищі. Фотосинтетичні та нефотосинтетичні мікроорганізми використовують світло як зовнішній стимул: при низьких рівнях освітлення вони демонструють позитивний фототаксис, прямуючи до водної поверхні з тим, щоб отримати достатнє для життєдіяльності сонячне випромінювання. Але при високих інтенсивностях сонячного випромінювання вони використовують негативний фототаксис, прямуючи у глибину з тим, щоб запобігти руйнуючій дії цього випромінювання. Фотоорієнтація не має місця у темряві, але є на значних глибинах, де світла не вистачає. В цьому разі водні організми використовують гравітацію як зовнішній фактор для вертикальної орієнтації.
Гравітаційне поле Землі є важливим зовнішнім фактором для організмів, що пересуваються у водному середовищі. Таку спроможність організмів орієнтувати напрямок свого руху відносно гравітаційного поля називають гравітаксисом. Гравітаксис спостерігається у таких водоростей, як Euglena gracilis, Chlamydomonas nivalis, Cryptomonas, Peridinium gatunense та P.faeroense, Amphidinium caterea, Prorocentrum micans, Dunalialla salina. Напрямок гравітаційної орієнтації організмів залежить від виду та віку водорості. Так, Peridinium faeroense та молода популяція Euglena gracilis виявляють позитивний гравітаксис (рух від водної поверхні), в той час як Chlamydomonas nivalis, Peridinium gatunense, Amphidinium caterea, Prorocentrum micans та старим популяціям Euglena gracilis притаманний негативний гравітаксис (рух до водної поверхні). Рівень гравітаційної орієнтації водоростей визначається віком культури та часом доби, наявністю важких металів у середовищі, впливом сонячного, зокрема, ультрафіолетового випромінювання. Таким чином, гравітаксис є важливим екологічним фактором, який дозволяє водним організмам знаходити кращі умови існування.
Щодо механізмів гравітаксису водоростей, то тут існує дві гіпотези. Згідно з першою гіпотезою пасивної переорієнтації, гравітаксис є чисто фізичним явищем, яке пояснюється нерівномірним розподілом маси в клітині [Brinkmann, 1968]. Важка частина клітини утворює обертаючий момент, який установлює клітину у вертикальне положення; завдяки дії джгутикового апарата Клітина прямує до водної поверхні. Але ця гіпотеза не підтверджується результатами останніх досліджень. Так, гравітаксис Euglena gracilis інгібується ультрафіолетовим випромінюванням, тоді як швидкість руху залишається незмінною [Hader and Liu, 1990]. Перша гіпотеза не спроможна пояснити, чому молодим популяціям Euglena gracilis притаманний позитивний гравітаксис, тоді як старим - негативний. Не пояснюється також реверсія гравітаксису під впливом важких металів. Експерименти, що були проведені у космосі, свідчать про те, що гравітаксис водоростей залежить від рівня гравітації, але не від магнітного чи електричного поля Землі. Отже, була запропонована друга гіпотеза [Lebert and Hader, 1996], згідно з якою гравітаксис є активний фізіологічний процес, в основі якого лежить різниця густини тіла клітини (1,05 г·мл-1) та середовища (1,00 г·мл-1), в якому вона знаходиться. Внаслідок седиментації тіло клітини утворює тиск на клітинну мембрану, що призводить до її зміщення на 1 нм та активізації чутливих до гравітації іонних каналів. На користь цієї гіпотези свідчать результати застосування специфічних препаратів - інгібітора механочутливих каналів гадолінію, іонофору А23187 та ванадату, які навіть при незначних концентраціях пригнічують гравітаксис, тоді як рухливість клітин залишається незмінною. Крім того, ці результати дають можливість допустити участь у механізмі гравіорієнтації електричних потенціалів, що виникають на цитоплазматичній мембрані. Типову схему спостереження та оцінки гравітаксису водоростей наведено на рис. 12.1.

Рис. 12.1. 1. Гравітаксис водоростей: а - типова схема спостереження та оцінки гравітаксис водоростей (1 - Мікроскоп; 2 - відеокамера, 3 - система обробки інформації; 4 - монітор спостереження за популяцію 5 - монітор спостереження за окремою клітиною; 6 - комп’ютер). б - гістограма кутового розподілу клітин зеленої водорост, Dunaniella seine лід час гравітаксису (кількість клітин 1336; середня швидкість 2536 мкм/с; рівень гравітаксису 0,352; напрямок гравітаксису 192,89°).
12.1.2. Гравітропізм рослин
Визначення гравітропаму. Ростову реакцію рослин, яка виклика вигин або скривлення її певної частини у бік зовнішнього стимулу, називають тропізмом. Якщо рух направлений до стимулу, має місце позитивний тропізм; якщо в протилежний бік - негативний тропізм. Коли зовнішнім стимулом є гравітаційна сила, то РЕАКЦІЯ РОСЛИНИ НА неї називається гравітропізмом. Рослина, покладена в горизонтальне положення (рис. 12.2), направляє паросток догори (негативний гравітропізм), а корінь - донизу (позитивний гравітропізм). Такі органи рослини, як еталони, ризоми, бокові гілки, що ростуть під прямим кутом до напрямку дії гравітаційної сили, демонструють діагравітропізм, а органи, напрямок росту яких знаходиться в інтервалі 0-90° до вертикалі, - плагіогравітропізм. Якщо органи не чутливі до гравітації, їх називають агравітропними. Типові гравітропні відгуки стебла та коріння рослин наведено на рис. 12.3.
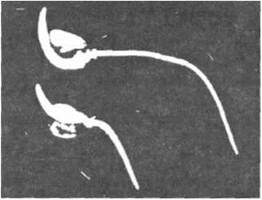
Рис. 12.2. Реакція рослини на гравітаційну силу: позитивний гравітропізм кореня та негативний — паростка.
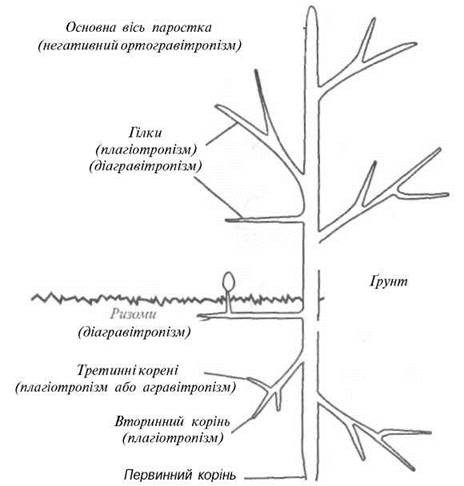
Рис. 12.3. Типові гравітропні відгуки стебла та коріння рослини.
Гравірецепція. На відміну від інших зовнішніх стимулів гравітація існує завжди і не змінюється за величиною. Крім того, гравітацію не можна увімкнути чи вимкнути; вона не характеризується наявністю градієнтів. Клітини на верхніх ділянках стебла чи кореня підлягають дії такої ж гравітаційної сили, що і на нижніх ділянках. Отже, такий зовнішній стимул, як гравітацію, можна реєструвати тільки завдяки руху певних рослинних чи клітинних структур - руху, внаслідок якого утворюється асиметрія в клітині, що призводить до виникнення тиску. У 1900 р. Хаберленд та Немек (цит. по [Weevers, 1949]) на основі ретельних цитологічних досліджень прийшли до висновку, що функції чутливих до гравітації органів у рослині виконують зерна крохмалю - так званих статолітів. Згідно з сучасними уявленнями механізм сприймання гравітаційної сили відбувається за рахунок осідання амілопластів - пластид, що містять зерна крохмалю (від 1 до 8) і розміщені у мембрані. Ці амілопласти, які мають певну масу і густину, рухаються по в'язкій цитоплазмі; при зміні положення рослини вони переміщуються донизу і затримуються біля клітинних стінок. Внаслідок свого накопичення вони створюють тиск на плазматичну мембрану або ендоплазматичний ретикулум (рис. 12.4). Рух рецепторів гравітації сприяє активації певних ферментів, що регулюють гормонний метаболізм. Внаслідок переміщення пластид утворюється градієнт ростових речовин, які забезпечують ріст паростка або кореня.

Рис. 12.4. Схематичне пояснення механізму гравірецепції. а - однорідний тиск статолітів на ендоплазматичний ретикулум при вертикальній орієнтації коріння; б - перерозподіл статолітів при горизонтальній орієнтації коріння, що обумовлює неоднорідний тиск статолітів на ендоплазматичний ретикулум ЕР.
Вимірювання гравітропізму. В більшості експериментів використовується гравітаційне прискорення на рівні 1 g чи менше, що досягається орієнтуванням органа (колеоптиля чи кореня) у положенні, що відрізняється від вертикального. Гравітаційна сила пропорційна синусу кута нахилу органа відносно вертикалі. Якщо потрібно звести нанівець вплив гравітаційного поля, застосовують кліностати - прилади, які забезпечують обертальний рух рослинного об'єкта, або проводять космічні досліди, в яких досягають стану невагомості.
12.1.3. Тигмонаетичні Рухи рослин
Настичні рухи виникають у відповідь на будь-які зовнішні фактори - Дотик, пошкодження, механічну вібрацію, світло, хімічні речовини, на які рослина реагує швидкими рухами. Напрямок настичних рухів не залежить від напрямку стимулу. Тигмонастичними називаються рухи, що викликаються дотиком; такі рухи характерні, наприклад, для добре відомої рослини мімоза соромлива (Mimosa pudica), яка згортає листки (а інколи - і все листя) протягом декількох секунд у відповідь на дотик. Такі швидкі рухи у мімози можуть бути використані для захисту проти комах та інших тварин, протидії суховіям з метою збереження води у посушливих районах, де вона мешкає. Зовнішній механічний стимул, що сприймається сенсорними клітинами черешка, перетворюється в електричний сигнал, який швидко поширюється по тканині, доки не досягне локомоторних клітин листкової подушечки поблизу основи листка, які негайно змінюють свій об'єм. Мабуть, рух органів у мімози визначається зміною тургорного тиску в цих клітинах. Зміна їхньго об'єму призводить до виділення з них іонів калію та таніноподібних речовин з клітини. Тигмонастичними можна вважати й рухи, які демонструють у відповідь на дотик Комахоїдні рослини - наприклад, венерина мухоловка Dionaea muscipula. Вважається, що швидке закривання листків рослини пов'язане з незворотним розтягуванням клітин та зміною підкислення клітинних стінок [Рейвн и др., 1990]. До тигмонастичних рухів слід віднести й закручування вусиків, за допомогою яких деякі рослини (Passiflora coerulea, Pisum sativum) здатні підтримувати себе у просторі. Вусики під час свого росту виконують кругові рухи з тим, щоб збільшити ймовірність контакту з потенційною опорою, внаслідок чого вони змінюють швидкість та напрямок росту так, щоб зачепитися за опору та підтягнути рослину. Якщо погладити нижню сторону вусика гороху Pisum sativum скляною паличкою, клітини нижньої сторони починають стискуватися, а клітини верхньої-розтягуватися, після чого всі клітини розтягуються, але клітини верхньої сторони розтягуються швидше. Очевидно, в регуляції зміни розмірів клітин беруть участь такі гормони, як ауксин, що стимулює ріст, та Етилен, який збільшує проникність мембрани; збільшення етилену в клітинах нижньої сторони супроводжується втратою розчинених речовин з вакуолі, що викликає стискування клітин.
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.