Фізіологія рослин - Мусієнко М.М. 2001
Фотосинтез: фізіолого-біохімічні та екологічні аспекти
Екологія фотосинтезу
Фотосинтетична продуктивність значно залежить від факторів довкілля — інтенсивності та спектрального складу світла, концентрації СО2, температури, водного режиму рослин, мінерального живлення, забруднення атмосфери шкідливими газами та важкими металами тощо. З дидактичних поглядів, кожен з названих факторів ми розглядаємо зокрема, тоді як в природі вони завжди діють в певному співвідношенні, взаємно. Тому будь-яку реакцію рослин слід розглядати як інтегральну відповідь з метою оптимізації системи через зміну окремих функцій.
Інтенсивність та якість світла.
Серед багатьох факторів, від яких залежить Фотосинтез, на перше місце слід поставити світло. Показником ефективності використання сонячної енергії є коефіцієнт корисної дії (ККД). ККД — це відношення кількості енергії, яка запасається в продуктах фотосинтезу або фітомасі врожаю, до кількості використаної сонячної радіації. Для різних рослин і в різних умовах вирощування цей коефіцієнт складає в середньому для кукурудзи 2,5-5,7; озимої пшениці 1,1-6,3; рису 2,5-4,4 тощо.
В середньому листки поглинають до 85% енергії фотосинтетично активних променів (400-700 нм) та до 25% енергії інфрачервоних променів, що становить близько 55% енергії загальної радіації. В той же час на фотосинтез витрачається лише 1,5-2% ФАР.
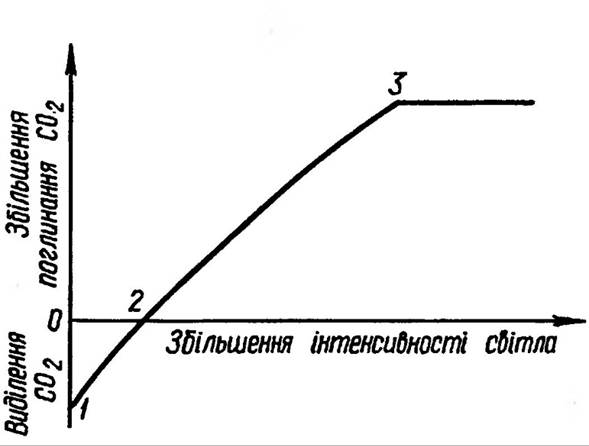
Рис 76. Світлова крива фотосинтезу
Узагальнену світлову криву фотосинтезу подано на рис. 76, де інтенсивність процесу розглядається як функція сили світла.
Початковий та середній ступені, які виражаються лінійною частиною кривої, не залежать від темнових реакцій. Та в міру зростання інтенсивності світла швидкість фотосинтетичних реакцій сповільнюється — спостерігається перегин світлової кривої і навіть вихід на плато При постійних величинах решти факторів для мезофітів полярних широт оптимум спостерігається при освітленні 20 000 лк, тоді як тіньовитривалі рослини часто уже при 2000-4000 лк досягають світлового насичення (рис. 77).
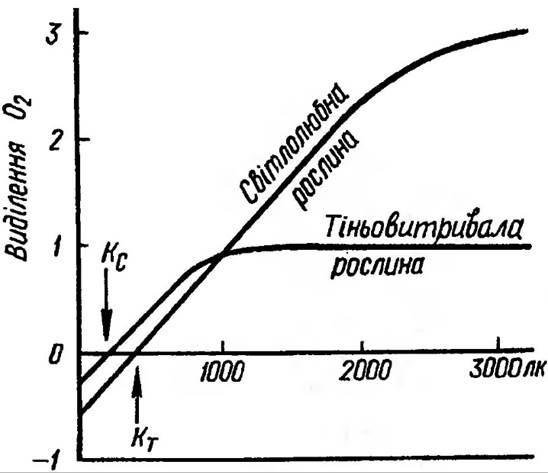
Рис 77. Залежність інтенсивності фотосинтезу від освітлення
Інтенсивність світла, при якій в міру зростання фотосинтезу компенсується темнове Дихання, називають точкою світлової компенсації, розміщення якої має особливо важливе значення для
продуктивності фотосинтезу. У тіньовитривалих рослин значення світлової компенсаційної точки становить до 1% від повного насичення, тоді як у світлолюбних — від 3 до 5%. Разом з підвищенням температури вона зміщується в бік більш високих значень освітленості і залежить від виду рослин, етапу онтогенезу та умов довкілля.
Різна інтенсивність світла, якісний склад його впливають і на утворення різних продуктів фотосинтезу. Наприклад, у листках квасолі слабка освітленість (до 2000 лк) сприяє синтезу переважно амінокислот і в меншій мірі — вуглеводів. Було виявлено, що сині промені спектра (458-480 нм), порівняно з червоними (670-680 нм) посилюють включення міченого вуглецю в малат, деякі амінокислоти та білки при одночасному зниженні його вмісту у вуглеводах. Виявлено також активацію синім світлом ФЕП-карбоксилази, яка відповідно впливає на включення 14С до складу С4-дикарбонових кислот. Характерно, що синє світло не впливає на активність рибулозобісфосфаткарбоксилази. Червоні промені виявились найефективнішими для фотосинтезу. Це пояснюється тим, що енергії кванта червоного світла (176 кДж/моль) цілком досить для фотозбудження молекули хлорофілу та переведення її на перший синглетний рівень, з якого і розпочинаються фотохімічні реакції. В цілому ж слід зазначити велику пластичність зеленої рослини по відношенню до зміни освітлення.
Вплив температури на фотосинтез, як правило, зворотній, якщо її значення не виходять за межі стійкості найважливіших систем листка. Також він залежить від інтенсивності освітлення. При низькій освітленості фотосинтез практично не залежить від температури, його інтенсивність однакова як при 15 °С так і при 25 °С. Це пояснюється тим, що при незначному освітленні інтенсивність фотосинтезу лімітується швидкістю протікання світлових реакцій. Навпаки, при високій освітленості швидкість фотоситезу залежить від швидкості світлових реакцій.
Майже для всіх рослин найсприятливішою є температура в діапазоні 10-35 °С, хоч досить часто пригнічення фотосинтезу спостерігається при порівняно невисоких температурах, вже після 25-30 °С. Деякі рослини пустелі здійснюють фотосинтез при 58 °С. Залежність швидкості фотосинтезу від температури для різних видів рослин подано на рисунку (рис 78). Мінімальна температура для фотосинтезу рослин середньої полоси близько 0 °С.
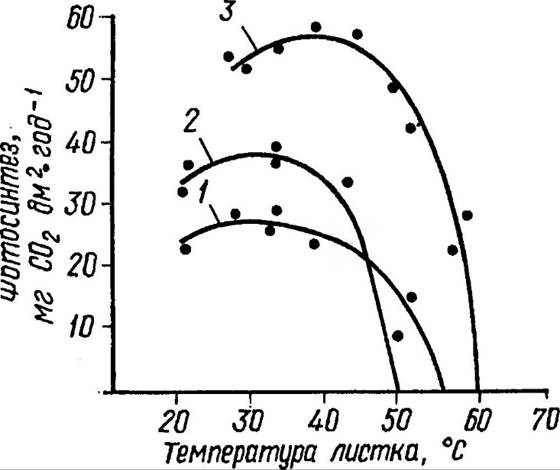
Рис 78. Вплив температури на інтенсивність фотосинтезу
Нижня температурна межа фотосинтезу для рослин північних широт — 15 °С, тоді як для рослин тропічного походження 4-8 °С. Однак верхні і нижні межі температури, а тим більше температурні оптимуми фотосинтезу, не можуть бути чітко фіксованими і залежать від виду, стадії розвитку та всього комплексу зовнішніх умов. Є дані, що фотосинтез тим вищий, чим менший температурний градієнт між рослиною і довкіллям та чим він більший між верхньою та нижньою частинами пластинки листка. Серед процесів фотосинтезу фіксація і
відновлення СО2 зазнає впливу температури швидше, ніж транспортування електронів по ЕТЛ. Найчутливіші до температури реакції карбоксилування, синтезу кінцевих продуктів відновлення СО2 — крохмалю та цукрози, а також транспортування фотоасимілятів до інших нефотосинтезуючих органів.
Слід пам’ятати, що поряд із прямим впливом на інтенсивність фотосинтезу температура як екологічний фактор впливає опосередковано на інші фізіологічні функції рослинного організму, тісно пов’язані з фотосинтетичним процесом.
Вплив вмісту СО2 в повітрі. В основному в процесі фотосинтезу використовується вуглекислий газ атмосферного повітря (0,03%), хоча є дані, що частково СО2 може надходити в рослину через кореневу систему із грунту.
Одним із обмежуючих факторів фотосинтезу є концентрація СО2 в повітрі, забезпечення рослин якою в природі залежить від впливу багатьох факторів: структури листка, щільності рослинного покриву, швидкості вітру тощо. На використання СО2 впливає опір певних ділянок фотосинтетичного апарата на шляху дифузії із повітря до ділянок фіксації СО2 (рис. 79).
Рослини С4-типу фотосинтезу (кукурудза, просо, сорго) володіють більш високою здатністю до засвоєння СО2 завдяки високій активності ферменту ФЕП-карбоксилази. Процес фотосинтезу може здійснюватися при вмісті СО2 для рослин С3-групи не менше 0,005%, тоді як для рослин С4-групи — 0, 0005%. Характерно, що навіть десятикратне збільшення концентрації СО2 в атмосфері спричинює наростання швидкості фотосинтезу. Насичення відбувається при концентрації 0,2-0,3% (рис. 80). При збільшенні вмісту вуглекислого газу до 15-20% процес фотосинтезу виходить на плато, а потім наступає депресія фотосинтезу. Високі концентрації
СO2 особливо несприятливі при високій освітленості.
Підвищена концентрація СО2 зумовлює синтез вуглеводів, аланіну, тоді як знижена — спричинює переважно утворення серину, гліцину та гліцеринової кислоти. Слід враховувати, що в денні години вміст СО2 в повітрі навколо рослин зменшується. Так, над полем кукурудзи він може знизитися майже втричі, до 0,01%. Це відбувається не дивлячись на те, що запаси СО2 поповнюються за рахунок переміщення шарів атмосферного повітря та виділення СО2 рослинами під час дихання.
Вплив водного режиму. Вода є необхідним безпосереднім учасником процесу фотосинтезу. Вона бере участь у фотосинтезі як субстрат окислення і як джерело кисню. Однак кількість води, яка необхідна для таких процесів, є мізерною в порівнянні із загальним вмістом води, необхідним для підтримування клітини в тургорному стані. Разом з тим, при повному насиченні водою клітин листка фотосинтез знижується, так як і при водному дефіциті вже від 15 до 20%.
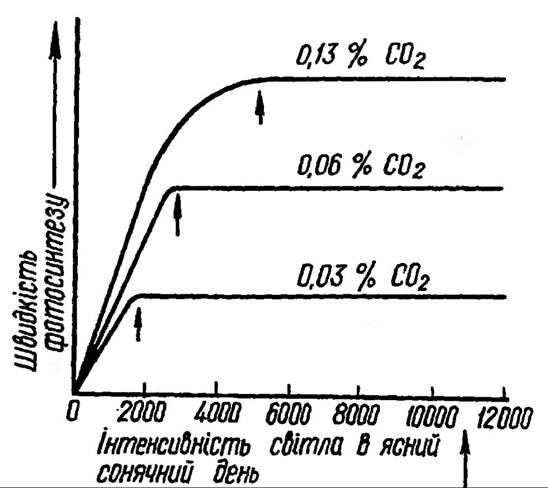
Рис. 79. Вплив концентрації СО2 на інтенсивність фотосинтезу
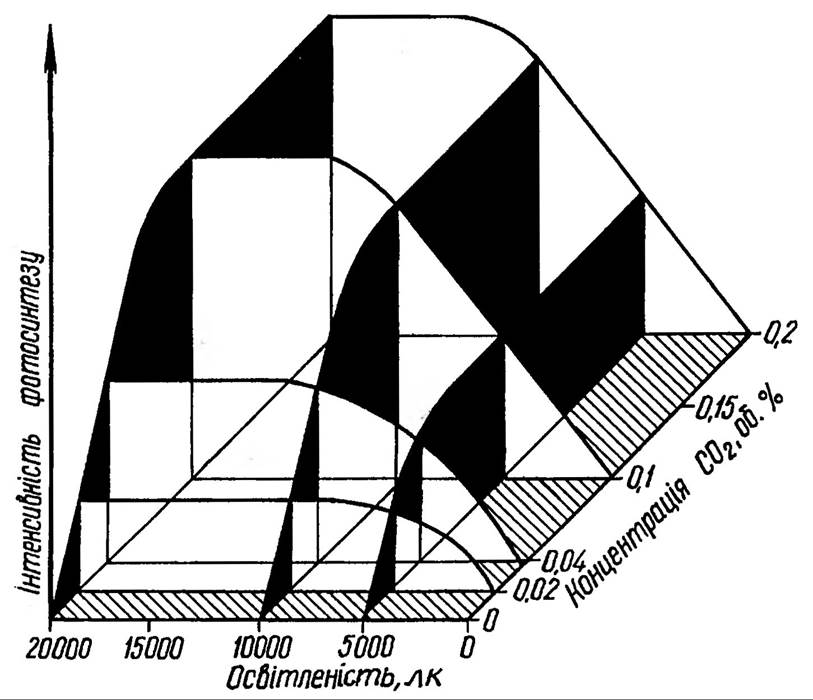
Рис 80. Залежність інтенсивності фотосинтезу від концентрацій СО2 та освітлення
Існуючий у природі компроміс між необхідністю підтримки високої швидкості надходження вуглецю та лімітуванням втрати води через продихи найтісніше зумовлений водним режимом, доступністю води для організму. Водний режим корелює з фотосинтезом насамперед через активність продихового апарата. Максимальний фотосинтез спостерігається при незначному водному дефіциті 5—20% від повного насичення при відкритих продихах. Дефіцит вологи впливає насамперед на темнову стадію, зокрема, на регенерацію первинного акцептора СО2 в циклі Кальвіна, тоді як електронне транспортування менш чутливе до зневоднення.
Вважають, що посуха діє неспецифічно — шляхом гальмування активності всіх розчинних пластидних та цитоплазматичних ферментів.
Якщо рослина перебуває в умовах посухи тривалий час, тоді змінюється її фотосинтетична продуктивність в цілому, зокрема, і через зменшення фотосинтетичної поверхні листків. Істотні втрати води врешті-решт спричинюють ультраструктурні зміни хлоропластів. При тривалому і значному дефіциті вологи процес фотосинтезу не відновлюється навіть після відновлення оптимального зволоження.
Вплив мінерального живлення. Одним із основних антропогенних факторів оптимізації умов життєдіяльності рослин, особливо в агрофітоценозах, з метою одержання максимальної продуктивності є регулювання рівня мінерального живлення. Вплив останнього проявляється не лише через інтенсифікацію роботи фотосинтетичного апарата, а й шляхом впливу на швидкість ростових процесів, від яких залежить площа листкової поверхні, густота продуктивного стеблостою. Елементи мінерального живлення потрібні для формування фотосинтетичного апарату, біосинтезу пігментів, як складові компоненти електрон-транспортного ланцюга, каталітичних систем хлоропластів.
В той же час більшість культурних рослин досить чутливі до підвищених концентрацій різних солей у грунті. Досить часто першочерговою відповідною реакцією на засолення є збільшення продихового опору по відношенню до дифузії СО2 Це пояснюється тим, що замикаючі клітини продихів втрачають здатність до Регулювання осмотичного тиску, бо надлишок іонів порушує функцію продихового апарату. Паралельно негативний вплив поширюється і на фотохімічні та біохімічні процеси. Застосування флуоресцентних зондів показало, що сольовий стрес впливає на функціонування світлозбираючих комплексів фотосистем.
Таким чином, реакція фотосинтетичного процесу на доступність певних мінеральних елементів свідчить про тісний взаємозв’язок між двома основними процесами живлення рослин — кореневим та повітряним.
Зміна фотосинтетичної активності рослинного організму є одним із найчутливіших показників забруднення атмосферного повітря. Атмосферне забруднення діоксидами сірки, азоту, вихлопними газами, які містять свинець, цинк, кадмій, нікель, талій, впливає насамперед на функціонування продихового апарату. Шкідливі концентрації сірчистого газу знижують величину квантового виходу фотосинтезу, інколи зумовлюють Набухання тилакоїдів, пригнічують потік електронів в межах другої фотосистеми.
Важкі метали в концентрації 0,5-10 м. д. (мільйонна доля) надзвичайно токсичні для рослинного організму. На жаль, більшість досліджень впливу важких металів на фотосинтетичні реакції проводилися in vitro, що не завжди адекватно поведінці рослинного організму in vivo. Тому щоб з’ясувати істинну картину пригнічення фотосинтезу забрудниками атмосферного повітря, слід вивчати їхній вплив на рослину в контролюючих умовах, зокрема в фітотроні.
Вплив внутрішніх факторів на інтенсивність фотосинтезу. Незважаючи на те, що процес фотосинтезу здійснюється в напівавтономних органелах — хлоропластах, він тісно поєднаний і контролюється певним чином іншими процесами, які відбуваються не лише в клітині, а і в організмі в цілому. Фотосинтез постачає первинні Органічні сполуки, а отже, впливає на накопичення маси рослини. В свою чергу, темпи росту і розвитку їх визначають інтенсивність використання продуктів фотосинтезу і тим самим впливають на швидкість цього процесу. Найважливішим фактором, який визначає інтенсивність процесу фотосинтезу, є донорно-акцепторні відносини. Донором виступає листок, а акцептором — різноманітні нефотосинтезуючі клітини та органи рослини.
Інтенсивність фотосинтезу зростає із збільшенням вмісту хлорофілу. Однак прямої про- порціональної залежності між цими двома показниками немає. За даними В.М.Любименко, із збільшенням вмісту хлорофілу асиміляційне число (кількість СО2 в мг, яка засвоюється одиницею вмісту хлорофілу за одиницю часу) знижується. Так, при збільшенні вмісту хлорофілу в 20 разів інтенсивність фотосинтезу збільшилася лише вдвічі, відповідно асиміляційне число знизилось в 10 разів. Оптимальним для фотосинтезу вважається вміст хлорофілу а від 3,1 до 5,7мг/дм2 незалежно від виду та віку рослин. В процесі Старіння листків інтенсивність фотосинтезу падає.
Денний хід фотосинтезу також неоднаковий. Фотосинтез розпочинається ранком, досягає максимуму в полудневі години, поступово знижується до вечора і припиняється із заходом Сонця. При підвищеній температурі, зниженні вологості максимум фотосинтезу зсувається на ранкові години. При напрузі метеорологічних факторів крива денного ходу фотосинтезу набуває двовершинний характер. В таких випадках в полудневі години фотосинтез різко знижується (явище полуденної депресії).
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.