Фізіологія рослин - Мусієнко М.М. 2001
Фотосинтез: фізіолого-біохімічні та екологічні аспекти
Міжклітинне паренхімне транспортування
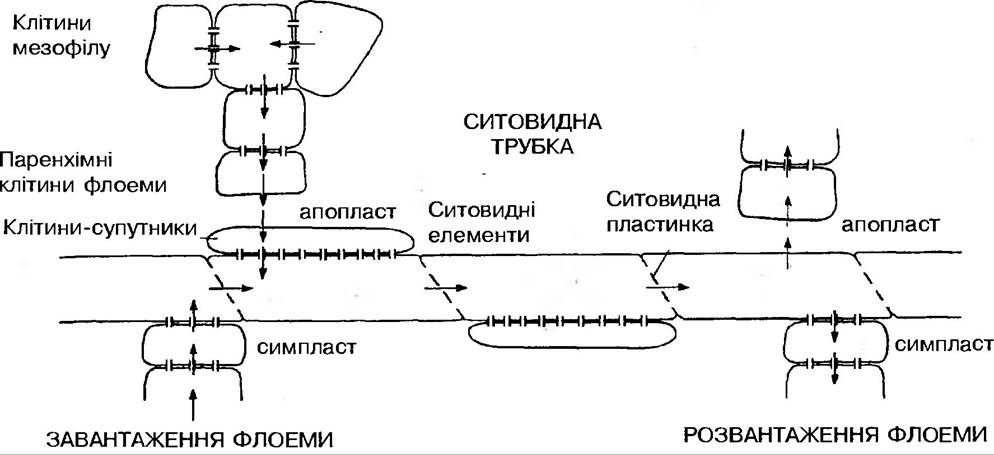
Пересування фотоасимілятів між клітинами паренхіми здійснюється двома шляхами — по симпласту через плазмодесми або за рахунок вільного внутріклітинного простору (найчастіше через клітинні оболонки паренхіми листка). Швидкість пересування асимілятів в паренхімних тканинах приблизно дорівнює 10-60 см за годину, що значно перевищує швидкість звичайної дифузії.
Встановлено, що клітини мезофілу листка мають значну секреторну здатність, тому легко виділяють цукри у міжклітинний простір. Передаточні клітини флоеми посилено адсорбують цукри та амінокислоти. Завдяки чисельним виростам клітинних стінок поверхня плазмалеми у передаточних клітин значно зростає, що створює оптимальні умови для поглинання асимілятів.
Вважають, що цукроза з клітин мезофілу листка десорбується і в апопласті:
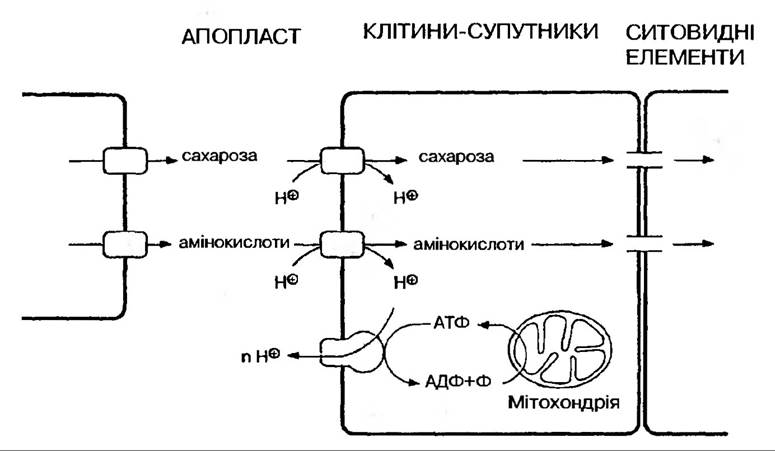
В міжклітинному просторі цукроза розщеплюється інвертазою до гексоз. Далі гексози пересуваються по апопласту до передаточних клітин провідних пучків за градієнтом концентрації. При першому дотику до паренхімних передаточних клітин флоеми вони знову перетворюються в цукрозу. Ймовірно, що частково цукроза із клітин мезофілу потрапляє до провідних пучків і через симпласт (по плазмодесмах). Далі відбувається завантаження ситовидних трубок і флоемний транспорт.
Флоемний транспорт фотоасимілятів. Вуглеводи, які синтезуються в листках під час фотосинтезу, транспортуються по судинах флоеми. Крім цукрів, в ситовидних трубках флоеми присутні амінокислоти, фітогормони та мінеральні речовини. На відміну від ксилеми — флоемний транспорт має вектор руху від донора, тобто місця, де синтезовані речовини, до акцептора, де вони асимілюються чи відкладаються про запас. Впродовж еволюції провідна система рослин поступово ускладнювалась. Так, у мохів для пересування асимілятів служать видовжені клітини. У водоростей поперечні стінки клітин вже перфоровані. Надалі, в процесі еволюції вже утворюються ситовидні трубки, які складаються із окремих члеників. Флоема являє собою сукупність живих клітин — власне ситовидні трубки, клітини-супутники, флоемна паренхіма та мертвих — луб’яні волокна. Ситовидні трубки являють собою вертикальні ряди витягнутих циліндричних клітин з тонкими клітинними оболонками, які формують неперервну систему. В місцях стикання клітин одна з одною їх стінки пронизані численними ситовидними порами. По мірі розвитку структура ситовидних трубок змінюється. Зникає Ядро, зменшуються розміри пластид, мітохондрій, зникає тонопласт. На місці вакуолі формується центральна порожнина, а Цитоплазма займає пристінне положення. Окремі повздовжні тяжі цитоплазми пронизують центральну порожнину. Одночасно в ситовидних пластинках утворюються пори, через які проходять тонкі тяжі цитоплазми. Інколи вони набувають форми мікротрубочок. Особливості будови клітин ситовидних трубок тісно пов’язані з їх функцією. До кожного членика ситовидної трубки примикає Клітина-супутник, яка має велике ядро, багато рибосом, мітохондрій, а також ендоплазматичний ретикулум. Основна їх функція полягає в забезпеченні енергією та інформаційною РНК.
Фотоасиміляти, синтезовані в листках, надходять в ситовидні трубки:
Пересування відбувається проти градієнта концентрації. Вважають, що проходить спільний транспорт цукрози і протону через специфічну пермеазу в плазмалемі ситовидних трубок. Такий котранспорт обумовлений протонним градієнтом. Градієнт протонів утворюється активно, тобто із витратами енергії, з допомогою протонної АТФази, яка постійно відкачує іони водню із ситовидних трубок. Витрати енергії вимагають часткового використання цукрози в ситовидних трубках. В постачанні енергією важливу роль відіграють суміжні клітини-супутники.
В ситовидних трубках відсутній фермент, який розкладає цукрозу (інвертаза), що і зумовлює цілісність даної сполуки на всьому шляху її транспорту. Транспорт по флоемі може відбуватись навіть одночасно в двох напрямках. Напрямок руху визначається виключно взаємним розміщенням донора і акцептора.
Поряд з транспортом фотоасимілятів із листків до кореня (низхідний потік) асиміляти можуть пересуватися вверх до точок росту, квітів, плодів. Допускають, що зустрічний потік речовин локалізований в різних ситовидних елементах. Швидкість пересування досить висока і досягає 50-100 см/годину. По флоемі в низхідному напрямку можна спостерігати пересування інших поживних речовин як у вигляді мінеральних, так і органічних сполук, наприклад, відтік із старіючих органів у процесі реутилізації.
Найбільш поширеною теорією, яка пояснює механізм транспорту асимілятів по флоемі, є теорія потоку під тиском. Вона узгоджується з наявними даними про градієнт тиску та осмотичний градієнт в ситовидних трубках на шляху транспорту.
Отже, якщо процеси завантаження флоеми та затримки цукрози в клітинах судинних пучків енергозалежні, то сам процес транспорту відбувається за чисто фізичними градієнтами.
В цілому швидкість транспорту залежить від швидкості утворення вуглеводів у донорних ділянках та швидкості використання в акцепторних, а тому залежить від метаболізму. Система донорно-акцепторних зв’язків забезпечує інтеграцію функціональних систем і цілісність рослинного організму. Донорно-акцепторні відношення регулюються гормональною системою та іншими ще мало дослідженими регуляторними факторами. Транспорт асимілятів забезпечує інтеграцію процесів фотосинтезу, росту та морфогенезу і є одним з головних факторів ендогенної регуляції фотосинтезу.
По мірі Старіння в порах ситовидних пластинок відкладається вуглевод калоза, яка звужує їх просвіт, що заважає рухові асимілятів. У деревних порід окремі елементи флоеми функціонують лише рік.
Значно впливають на пересування фотоасимілятів умови довкілля, насамперед Температура, а також умови мінерального живлення. Фосфор, бор помітно прискорюють їх транспорт. Є дані, що іони калію підтримують мембранний потенціал в ситовидних пластинках, що і сприяє прискореному транспорту. Існує тісний взаємозв’язок між пересуванням фотоасимілятів та загальним метаболізмом рослинного організму.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.