Фізіологія рослин - Мусієнко М.М. 2001
Водний режим рослин
Коренева система як орган поглинання води
Для виконання своїх основних функцій — поглинання із грунту води та мінеральних солей — корінь повинен володіти здатністю орієнтуватися в просторі, реагувати на градієнти життєво важливих факторів, просуватися у відповідності з цим в грунтових профілях і створювати максимальну поверхню для контакту з грунтом.
Всі особливості будови кореня найтісніше пов’язані з виконанням таких функцій. Роль даного органу полягає насамперед у тому, що завдяки його величезній поверхні забезпечується надходження води в рослину із великого об’єму грунту. Так, у однієї рослини озимого жита за сприятливих умов може утворитися 143 корені першого порядку, 35 тисяч — другого, 2 млн. 300 тис. — третього і 11,5 млн. — четвертого порядку, разом 14 млн. коренів. Загальна довжина їх складає 600 км, загальна поверхня 225 м2. На них розміщені 15 млрд. кореневих волосків, загальна довжина яких біля 10 тис. км, а площа 400 м2. Разом з тим надземна частина цієї ж рослини жита (всі пагони та листки) складає загальну площу приблизно 4,5 м2 (Лебедев, 1978). Коренева система здатна активно пересувати воду в клітинах кореня в певному напрямку — через кореневі волоски, клітини корової паренхіми, ендодерму, перицикл до судин ксилеми осьового циліндра кореня (рис 25).
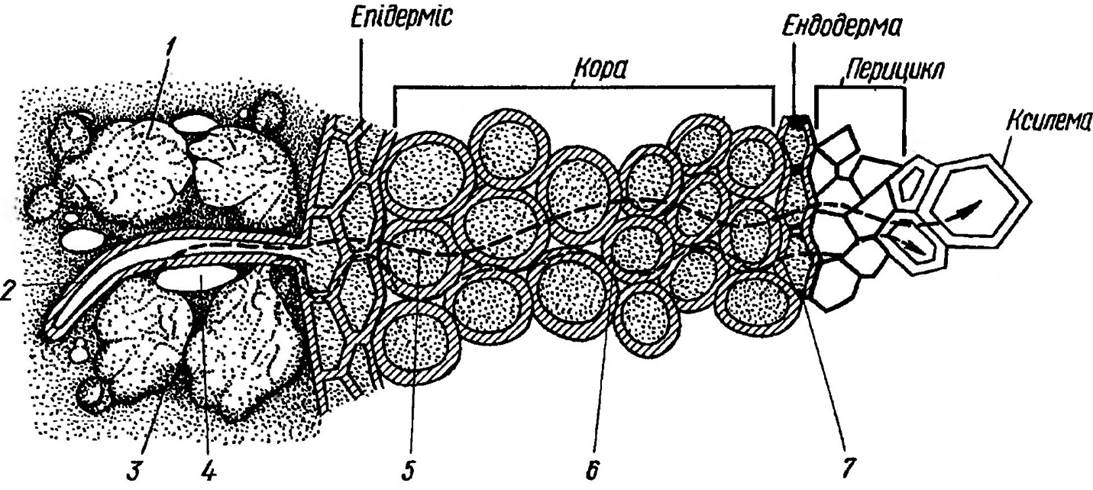
Рис 25. Надходження води з грунту в клітини кореня:
1 — грунтові частинки, 2 — кореневий волосок. З — ґрунтова Вода, 4 — повітряний простір, 5 — симпласт, 6 — апопласт, 7 — поясок Каспарі
Зона найбільш інтенсивного поглинання води коренем співпадає із його зоною розтягування та зоною розвитку кореневих волосків. Деяка кількість води може надходити і через опробковілу зону кореня. Це характерно головним чином для дерев. В таких випадках вода проникає через сочевички або поранення кореня. Через клітини паренхіми кори можливі два шляхи транспорту води — через цитоплазму по плазмодесмах (симпласт) і через клітинні стінки (апопласт). Оскільки опір клітинних стінок для води значно нижчий, ніж у цитоплазмі, радіальний транспорт води здійснюється в цій частині кореня, в основному, апопластним шляхом. Однак на рівні ендодерми такий шлях пересування води стає неможливим із-за непроникних для води поясків Каспарі в її клітинних стінках.
Вода в цій частині кореня може подолати ендодерму лише пройшовши через мембрани і цитоплазму цих клітин. Регуляція надходження води на рівні ендодерми здійснюється, з одного боку через зміну швидкого апопластного пересування води на більш повільний симпласт- ний, а з іншого — тим, що діаметр осьового циліндру кореня, куди в подальшому надходить
вода через ендодерму, в 5-6 разів менше діаметра поверхні кори та всисної зони кореня. Слід відмітити, що непроникність клітинних стінок ендодерми для води не є абсолютною. Зокрема в ендодермі тих ділянок кореня, які продовжують свій ріст, пояски Каспарі ще повністю не сформовані, а тому і зміни типів транспортування води, ймовірно, не буває. Крім того, в ділянках кореня, де закладаються бічні корені, ендодерма переривається і вода може продовжити свій шлях через пропускні клітини. Однак, в цілому масовий потік води через ендодерму по апопласту різко знижується. Всередині осьового циліндру рух води від ендодерми до судин ксилеми зустрічає незначний опір і, ймовірно, також здійснюється апопластним шляхом.
Два основні процеси забезпечують транспортування води та розчинених в ній речовин по рослині: транспіраційний потік та пересування фотоасимілятів (рис. 26). Для такого транспортування існують Провідні Тканини: судини та трахеїди в ксилемі для транспіраційного потоку і ситовидні трубки флоеми для асимілятів. Це шляхи дальнього транспортування.
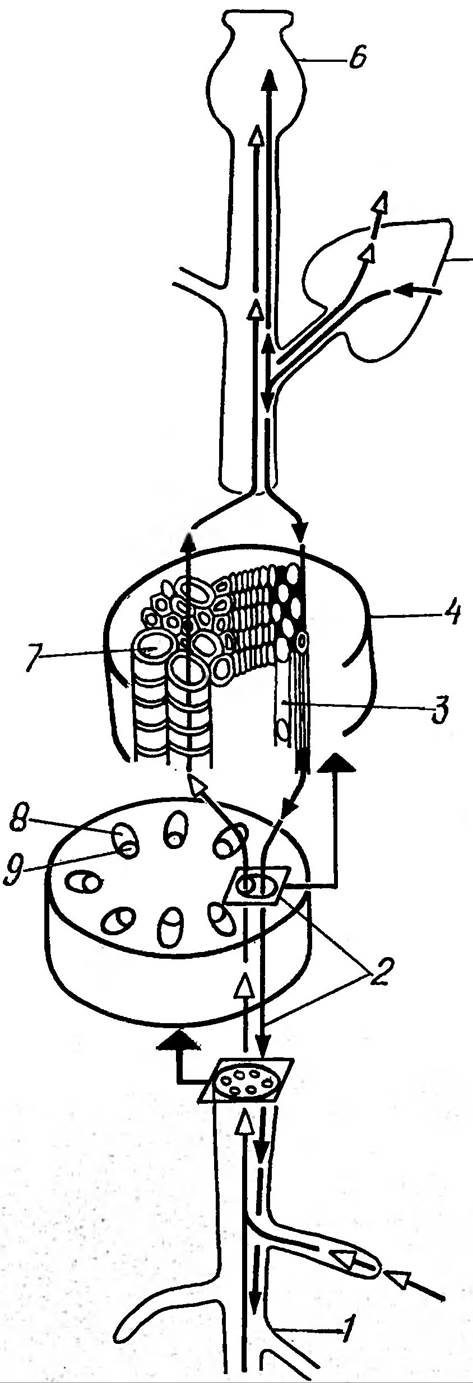
Рис. 26. Схема зображення головних транспортний шляхів транспіраційного потоку ( — — Δ) та потоку асимілятів (— — →):
1 — коріння, 2 — вісь пагона, 3 — ситовидна трубка, 4 — провідний пучок, 5 — листок, 6 — плід; 7 судина, 8 — флоема, 9 — ксилема
Між флоемою та ксилемою існують клітини- транслокатори, функція яких — переключати потоки флоемного та ксилемного транспортування. Ці клітини мають специфічні білки — транслокатори, молекулярний Механізм дії яких поки що невідомий.
Ближнє транспортування до провідних судин та від них здійснюється через клітини неспеціалізованих тканин. Останні складаються з цілого ряду розмежованих, тобто відокремлених одна від одної систем, — апопласту та симпласту.
Всі ці системи служать для ближнього транспортування: симпласт — для мінеральних та органічних речовин, апопласт — для води та неорганічних солей. Вакуолі як дискретні системи служать виключно для води.
Слід зауважити, що транспортування по симпласту відбувається швидше, ніж дифузія (1-6 см за годину). Механізм руху не вивчений, можливо в ньому бере участь дифузія, а також конвекція (перемішування речовин за участю ендоплазматичного ретикулуму).
Вода пересувається по рослині із зони з високим водним потенціалом (грунт) в зону з більш низьким водним потенціалом (атмосфера) відповідно з його градієнтом. Водний потенціал помірно зволоженого повітря на кілька десятків тисяч кілопаскалей нижчий ніж в рослині, тому вода і прагне покинути тканину рослин.
Сили висхідного потоку води в рослині. Загальновизнано, що поглинання води та пересування її вверх здійснюється в результаті спільної дії таких факторів, як кореневий тиск або нижній кінцевий двигун (рис. 27) та Транспірація — Верхній кінцевий двигун (рис. 28). Вода, яка надійшла в клітини кореня, під впливом різниці водних потенціалів, які виникають завдяки транспірації і кореневому тиску, пересувається провідними елементами ксилеми. В нормі вода із рослини виходить у вигляді пари. Для переходу води з рідкого стану в пароподібний використовується сонячна енергія. Випаровується вода че
рез продихи, кутикулу та сочевички (у листопадних дерев після опадання листя).
Отже, в природних умовах у ксилемі існує потік води як результат транспірації. При слабкій транспірації концентрація солей в ксилемі зростає і за законом осмосу сприяє руху всередину кореня. Вода, рухаючись через тканини кореня до центрального циліндра, в клітинах ендодерми повинна проникнути крізь їхню мембрану та протопласти, оскільки їхні стінки непроникні

Рис. 27. Схема, що пояснює як під впливом пересування води з кореня в стебло створюється кореневий тиск:
1 — манометрична трубка, 2 — ртуть,
З — пасока, 4 — гумове з'єднання, 5 — пеньок зрізаного пагона
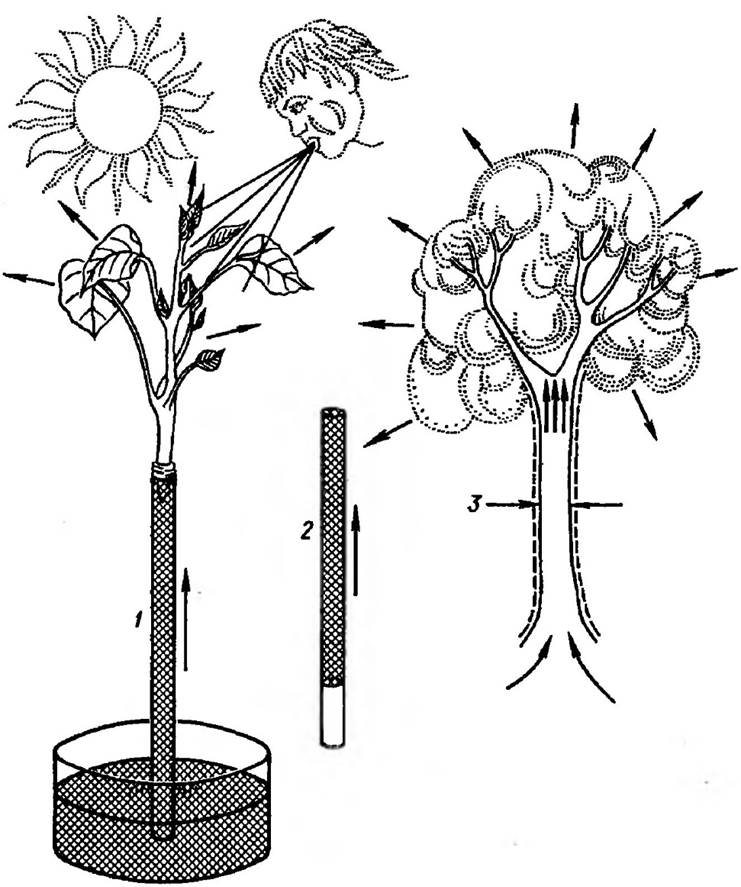
Рис. 28. Механізм підняття води в рослинах:
1 — в умовах Інтенсивної транспірації пагін може забезпечувати підняття води на значну висоту, яка перевищує барометричну; 2 — водяний стовпчик, який не містить газу, піднімається як єдине ціле завдяки силі зчеплення між молекулами води; 3 — коли втрата води перевищує її поглинання, в судинах виникає натягнення води і стінки судин зтягуються.
для води. Тому циліндричний прошарок клітин ендодерми є немов єдиною мембраною, по один бік якої (в ксилемі) знаходиться концентрований розчин, а по інший (з боку грунту та тканин кореня) — розчин значно слабшої концентрації. Саме тому вода дифундує з грунту в ксилему крізь цю «так звану мембрану» внаслідок різниці концентрацій.
Непроникні клітини ендодерми (правильніше стінки їх) виконують ще одну функцію; вони не дають змоги для солей, що надійшли в ксилему, здійснити зворотний шлях, тобто вийти назовні крізь апопласт. Ось в таких умовах в кореневій системі розвивається тиск в кілька атмосфер — кореневий тиск. При основі стовбура дерев він досягає 1,013 МПа. Якщо перерізати стебло трав’янистої рослини недалеко від грунту, то із зони надрізу починає виділитися пасока. Це явище називають плачем рослин.
Плач рослин доводить наявність кореневого тиску. Останній можна виміряти за допомогою звичайного ртутного манометра як це показано на рис. 28. Його величина навіть у трав’янистих рослин досягає 2-3 атм. Виникає запитання, а чи не є плач рослин патологічним явищем, що виникає внаслідок поранення рослини при зрізі?
Про те, що плач рослин є нормальним явищем, свідчить інше явище — гутація. Гутація — виділення краплин води крізь гідатоди — особливі пори по краях листків в місцях закінчення листкових жилок. Гутація в природніх умовах відбувається, головним чином вранці, коли кореневий тиск зростає. Гутації також сприяє помірно тепла та волога погода, коли оточуюче рослину повітря насичене водяним паром (рис. 29). В лабораторії її легко спостерігати, закривши, наприклад, проростки пшениці скляним ковпаком. На кінчиках молодих листочків пшениці будуть виділятися краплини води. Під час гутації ніякого пошкодження рослин не відбувається, однак, завдяки наявності кореневого тиску, рослина виділяє краплини води. Причому в рослин є спеціальні пристосування для захисту від можливих втрат мінеральних речовин при гутації. Справа в тому, що гідатоди виділяють воду в повітряну порожнину, вистелену дрібними клітинами паренхіми, так званою епітемою, фільтруючись крізь яку вода залишає в ній більшість мінеральних речовин (рис. 30).
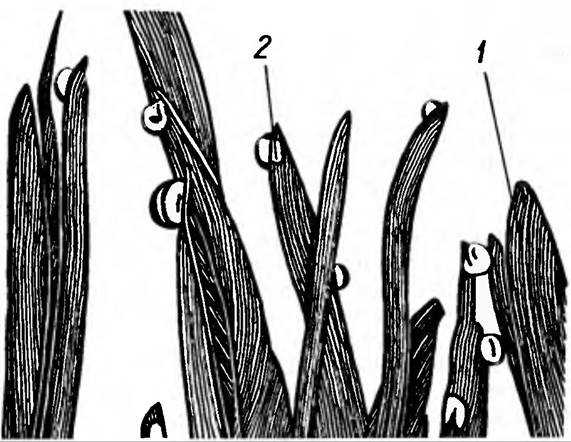
Рис. 29. Гутація.
Краплини води (2) на кінчиках листочків (1) свідчать про наявність кореневого тиску. Ці краплини не конденсуються з водяної пари оточуючого повітря. Вони витискуються з листка кріз спеціальні отвори — гідатоди, які знаходяться на його кінчиках
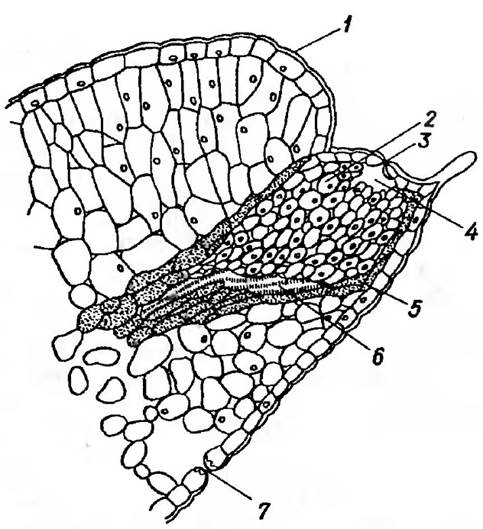
Рис. 30. Поздовжній розріз через гідатоду листка:
1 — кутикула, 2 — епітема, 3 — водний продих (пора), 4 — субепідермальна порожнина, 5 — обкладинка, 6 — трахеїда, 7 — продихова щілина
Отже, плач рослин, як і гутація показують, що коренева система поглинає воду і накачує її в рослину. На цій основі кореневу систему рослин називають нижнім кінцевим двигуном.
Силу, яка спричинює в рослині односторонній потік води з розчиненими речовинами незалежно від транспірації, називають кореневим тиском. Характерно, що не лише змертвіння клітин кореня, а і зниження їх життєдіяльності, зокрема, інтенсивності Дихання, припиняє плач рослин.
За Д.А. Сабініним плач рослин — це прижиттєвий односторонній потік води, пов’язаний із витратами енергії. Механізм даного явища повністю не з’ясовано. Допускають, що вода пересувається пасивно осмотичним шляхом за градієнтом водного потенціалу. Більш від’ємний, тобто низький водний потенціал, створюється в судинах ксилеми завдяки надходженню туди із грунту розчинних солей та через слабку протидію нееластичних її стінок. Солі надходять активно, для чого і потрібна енергія. Тому кореневий тиск можна визначити як тиск, що розвивається в ксилемі завдяки метаболічній активності кореня. Так як енергія постачається за рахунок дихання, робота нижнього кінцевого двигуна — кореневого тиску, корелює з наявністю кисню та залежить від присутності інгибіторів та активаторів процесу дихання. За даними В.М. Жолкевича кореневий тиск складається із двох складових частин: осмотичної та метаболічної, причому остання потребує затрат АТФ. Передбачають, що при цьому значну роль відіграють актиноподібні білки, енергозалежне скорочення та розслаблення яких і спричинює зміни гідростатичного тиску в клітинах. Внаслідок цього на шляху водного потоку в напрямку судин ксилеми виникають локальні градієнти водного потенціалу, які і сприяють надходженню води в судини.
Останнім часом з’явилися дані про наявність так званих водяних помп в коренях рослин (Кундт, Робник,1998). Такими помпами можуть бути скоротливі клітини, які мають клапани в плазмодесмах. Передбачають, що вони локалізовані в ендодермі, інколи в екзодермі. Стінки таких помпукріплені хвилястими поясками Каспарі, а роль клапанів виконують розміщені в зовнішніх периклинальних клітинних стінках перові поля, кожне з яких пронизане великою кількістю плазмодесм. Субмікронні складки зовнішніх периклинальних стінок виконують роль рухомого поршня під час роботи помп. На думку цих авторів, кореневий тиск створюється зусиллями багатьох клітин помп, що працюють з частотою близько 1 Гц, тобто подібно до серця людини.
У деяких багаторічних рослин взимку судини ксилеми заповнені повітрям, тому весною надходження води здійснюється лише за рахунок нижнього кінцевого двигуна.
Згадаймо також рух пасоки, який не має прямого відношення до кореневого тиску. Витікання соку весною зумовлене насамперед появою тиску в стовбурі дерев. Сонячні теплі дні, особливо коли вони чергуються з холодними ночами, спричинюють інтенсивний потік пасоки. Справа в тому, що в холодні ночі відкладений в паренхімних клітинах ксилеми крохмаль гідролізується, перетворюючись на цукри, які потім інтенсивно транспортуються по судинах ксилеми. З підвищенням температури з розчину виділяється СО2, в результаті чого в ксилемі виникає тиск. Саме цей тиск і змушує воду разом з розчиненими в ній цукрами пересуватися вгору по стовбуру дерева перед початком весняного росту. Доказом вище зазначеного може бути приклад із зрубаним молодим деревцем: якщо зрубане деревце помістити в бочку з водою, то при виділенні соку вода буде відсмоктуватись з бочки, незважаючи на відсутність кореневої системи.
Таким чином вода, що надійшла в клітини кореня, під впливом різниці водних потенціалів, а можливо і за участю водяних помп, рухається до провідних елементів ксилеми, щоб розпочати її висхідний рух до надземних органів. Приблизний розподіл водних потенціалів, які забезпечують пересування води, такий: водний потенціал грунту (-0,05 МПа), кореня (-0,2 МПа), стебла (-0,5 МПа), листків (-1,5 МПа), повітря при відносній вологості 50% (-100,0 МПа). Досліди з окільцьовуванням показали, що висхідний потік води по рослині здійснюється, в основному по ксилемі. Лише незначна її частина рухається поза ксилемою.
Швидкість пересування води визначається різницею водних потенціалів на початку і в кінці шляху, а також опором, який вода зустрічає.
Фізіологічна роль стебла. Підйом води в стовбурах дерев. Подана коренем вода швидко пересувається по рослині до листків. Дерева здатні подавати воду із грунту в крону в кількостях, часто перевершуючи одну тону в день. Яким чином це відбувається? Поглинена кореневими волосками вода проходить відстань в декілька міліметрів живими клітинами і лише потім уже надходить в мертві судини ксилеми. Пересування води живими клітинами можливе завдяки різниці їх водних потенціалів. Всисна сила зростає від кореневого волоска до тих живих клітин, що примикають до судин ксилеми. Встановлено, що для пересування води живими клітинами, наприклад листка, всисна сила кожної наступної клітини повинна відрізнятися на 0,1 атмосфери.
Звідси стає зрозумілим, чому рослини, які не мають судин (мохи, Лишайники), не досягають великих розмірів. Лише з появою трахеїд (папоротеподібні, голонасінні) та судин (покритонасінні) в процесі еволюції створилась можливість для рослин досягати висоти в декілька десятків і навіть більше сотні (евкаліпти, секвої) метрів. Тільки невелику частину свого шляху в рослині вода проходить по живих клітинах — в коренях, а потім в листках. Випаровування води з поверхні листків створює всисну силу в клітинах листків і коренів, чим підтримує пересування води по рослині. Тому листки рослин і отримали назву верхнього кінцевого двигуна, на відміну від кореневої системи, яка нагнітає воду в рослину. Випаровуючи воду з поверхні своїх листків, рослини автоматично втягують воду судинами.
В той же час відомо, що будь який всисний насос не може підняти воду на висоту, що перевищує 10 м, так як вага такого водяного стовпа буде відповідати атмосферному тиску, а отже ним зрівноважуватися. Різниця між таким насосом і стеблом обумовлена силами зчеплення води із стінками судин та між власними молекулами. Суцільні водяні стовпчики, що заповнюють судини, міцно спаяні завдяки силі зчеплення. Вага стовпчика води в 100 м висоти відповідає всього 10 атмосферам. Така величезна сила зчеплення дозволяє воді в стеблах рослин підніматися на висоту, яка значно перевищує барометричну.
Кореневий тиск та присисна дія листків рухають водяний потік на значну висоту. Велике значення при цьому мають також поперечні перегородки в судинах, так як повітря, попадаючи в судини, ізолюється, і із загальної системи водопостачання виключаються лише незначні ділянки.
Нагадуємо, що водний потенціал помірно зволоженого атмосферного повітря становить від’ємну величину і є на кілька десятків тисяч кілопаскалей нижчий ніж в рослині, тому вода і прагне покинути її. За рахунок транспірації водний потенціал у верхівці рослини нижчий, ніж при основі його. Крім того, швидкість випаровування води більша швидкості її надходжження. Тому в водних стовпчиках, які заповнюють ксилему високих дерев, існує від’ємний тиск, тобто натяг. Справа в тому, що в судинах ксилеми молекули води зв’язані не тільки одна з одною за рахунок сил зчеплення — когезії, — а і з стінками судин, які дуже гідрофільні, за рахунок сил прилипання — адгезії. Опір стовпчика води до розривання становить 30 МПа, цього досить, щоб підняти воду на висоту 130-140 м.
Ці сили не дають змоги розірвати водні тяжі й утворити пустоти — порожнини. Саме спільність транспіраційного потоку, капілярних, осмотичних сил, а також когезії та адгезії і зумовлює піднімання води в стовбурах дерев. Допускають участь в цих процесах і водяних помп в коренях рослин. Швидкість руху води судинами порівняльно невелика. Для листяних деревних порід вона складає в середньому 20 см3 за годину на 1 см2 поперечного розрізу деревини, а для хвойних всього 5 см3 за годину. Для порівняння, Кров артеріями рухається із швидкістю 40-50 см3 за секунду, а вода водопровідними магістралями 100 см3 на 1 см2 перетину в секунду.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.