Фізіологія рослин - Мусієнко М.М. 2001
Вплив зовнішніх умов довкілля на ріст і розвиток рослин. Фотоморфогенез. Фітохром. Фотоперіодизм. Яровизація
Регуляція росту і розвитку рослин світлом
Вплив зовнішніх умов на РІСТ І РОЗВИТОК рослин обумовлений різноманітними чинниками, серед яких можна виділити: кліматичні (Вода, світло, тепло, газовий склад повітря тощо); едафічні (структура грунту, його хімічний склад); біотичні (різноманітні мікроорганізми, рослини, тварини). Всі вони діють в сукупності, і їх дія найтісніше пов’ язана між собою та взаємно- обумовлена. У попередніх розділах було розглянуто роль води, грунту, мінерального та повітряного живлення у життєдіяльності рослинних організмів, тому в даній главі ми зупинимось на таких важливих чинниках довкілля як світло і Температура.
Багато ростових процесів, таких як швидкість росту, напрямок росту і процеси розвитку — цвітіння, плодоутворення, листопад, перехід до стану спокою, Старіння — залежать від світла, регулюються ним. Однак в цих випадках головна роль належить не інтенсивності світла, а, насамперед, тривалості періодів освітлення і темряви. Адже, крім забезпечення процесів фотосинтезу, світло виконує сигнальну та регуляторну функції. Зелена рослина — це не просто агрегат, який працює на сонячній енергії. Сама структура цього механізму частково визначається кількістю та якістю світлової енергії, яку вона поглинає.
Найкраще можна продемонструвати роль світла при вирощуванні рослини в темряві;
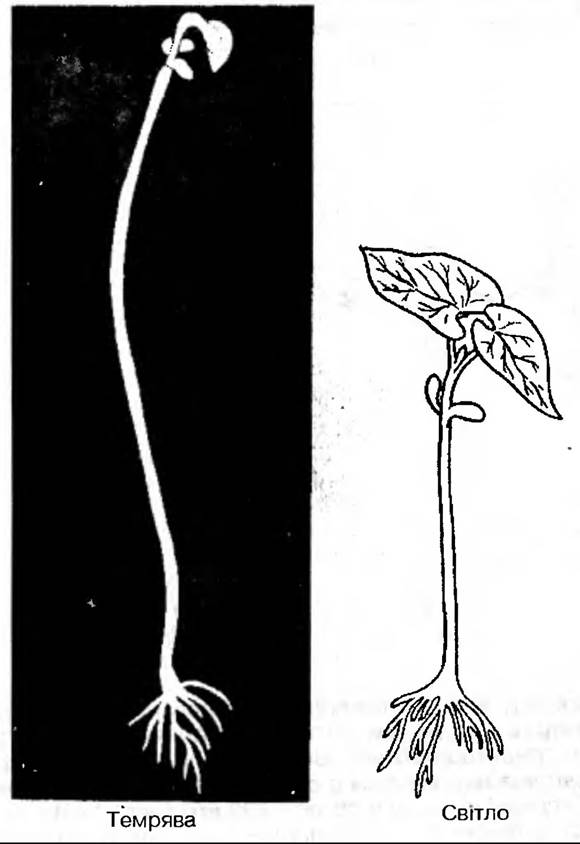
В таких рослин не нагромаджується хлорофіл, міжвузля видовжені і тонкі, листочки залишаються дрібними, недорозвинутими. Замість мембранної системи хлоропластів в них утворюються етіопласти. У таких рослин мало опорних тканин, тому вони легко ламаються, полягають. Рослини, вирощені в темряві, називають етіольованими, а саме явище — етіоляцією. Природна етіоляція має і позитивне значення, бо вона дозволяє рослині максимально рости в довжину з мінімальною затратою вуглецевих запасів, які не відновлюються без фотосинтезу. Якщо етіольований проросток не вийде на світло і не почне фотосинтезувати, то після витрати всіх запасних поживних речовин він приречений на загибель.
На проросток, вирощений в темряві, істотно впливають навіть незначні дози поглинених сонячних квантів: змінюється швидкість та напрямок росту листків і стебел, опушення, пігментація та ін.
Від освітлення залежить перехід до цвітіння та плодоутворення. Нарешті, опадання листя, перехід до стану спокою, феномен старіння — це все наслідок дії раніше поглинутих світлових та інших сигналів. Всю різноманітність процесів, пов’язаних із зпливом світла на ріст та диференціювання рослин, називають фотоморфогенезом. У процесі фотоморфогенезу відповідно до генотипу рослина набуває форми, оптимальної для конкретних умов. Іншими словами, фото- морфогенез — це нормальний ріст і розвиток рослини. Причому, Фотосинтетичні пігменти не відіграють в цих процесах головної ролі, а основну функцію в них виконують фікобілінові фітохромні пігменти та деякі інші, можливо, флавопротеїдні пігменти.
Фітохром
Механізм дії світла на морфогенетичні реакції ще мало досліджений. При фотоморфогенезі світловий сигнал ініціює експресію нової генетичної програми — створення зеленої рослини. Фоторецептори — фітохром та криптохром індукують всі реакції фотоморфогенезу. Вміст фітохрому в рослині незначний, тому не дивно, що кількість енергії, яка потрібна для насичення відповідних фотопроцесів, на кілька порядків нижча, ніж при фотосинтезі. Продукти цих світлових реакцій не регулюють ріст рослин стехіометрично, так як це буває з вуглеводами при фотосинтезі. Скоріше вони впливають на такі параметри, як проникність мембран, функція генів та активність ферментів. Тому для порівняння можна сказати, що коли при фотосинтезі на виділення одного моля кисню слід витратити 8-10 квантів, то така ж кількість квантів, поглинутих фітохромною системою, може повністю визначити параметри росту всього стебла, формування репродуктивного періоду.
Фітохром — це розчинний у воді хромопротеїд, хромофор якого є тетрапірол з незамкненим ланцюгом, який тіоефірним зв'язком з'єднується з апопротеїном:
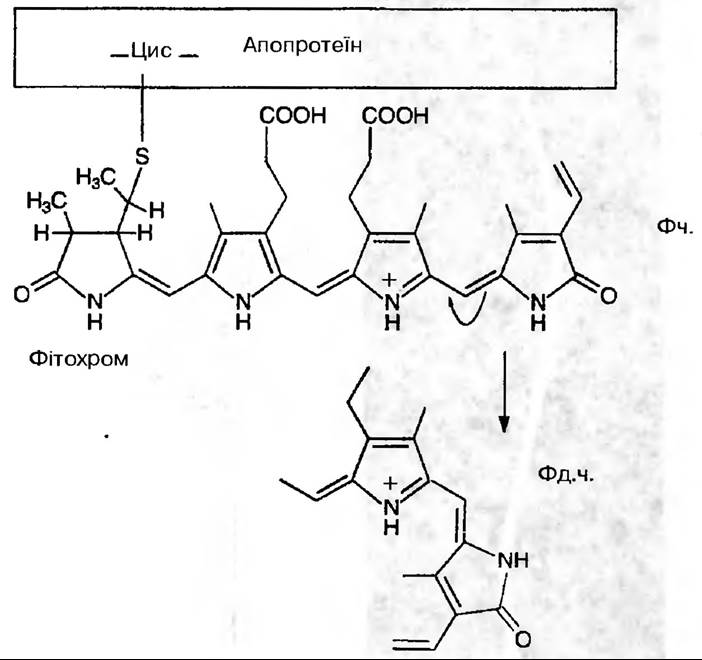
Це біліпротеїновий пігмент, який, ймовірно, локалізований в мембранах рослини і може легко змінювати свою форму та максимуми світлопоглинання під дією світла. Фітохром має дві форми, які постійно взаємноперетворюються. Вони дістали назву Фч. (фітохром червоний, який поглинає червоні промені світла з максимумом в області 660 нм) та Фд.ч. (фітохром далекий червоний, який поглинає дальні червоні промені в області 730 нм) (рис. 186).
Фітохром синтезується у формі Фч, фізіологічно неактивній. При опроміненні червоним світлом більша частина його переходить у форму Фд.ч. — фізіологічно активну форму. Форма Фд.ч. досить лабільна, тому в темряві вона поступово знову перетворюється на Фч. (Фч -> Фд.ч.→ Фч). Отже, фітохром функціонує як біологічний вимикач. В будь-яких випадках ефект червоного імпульсу на відновлення високого рівня Фд.ч. може бути знятий коротким опроміненням далеким червоним світлом, яке переводить знову Фд.ч.→ Фч .
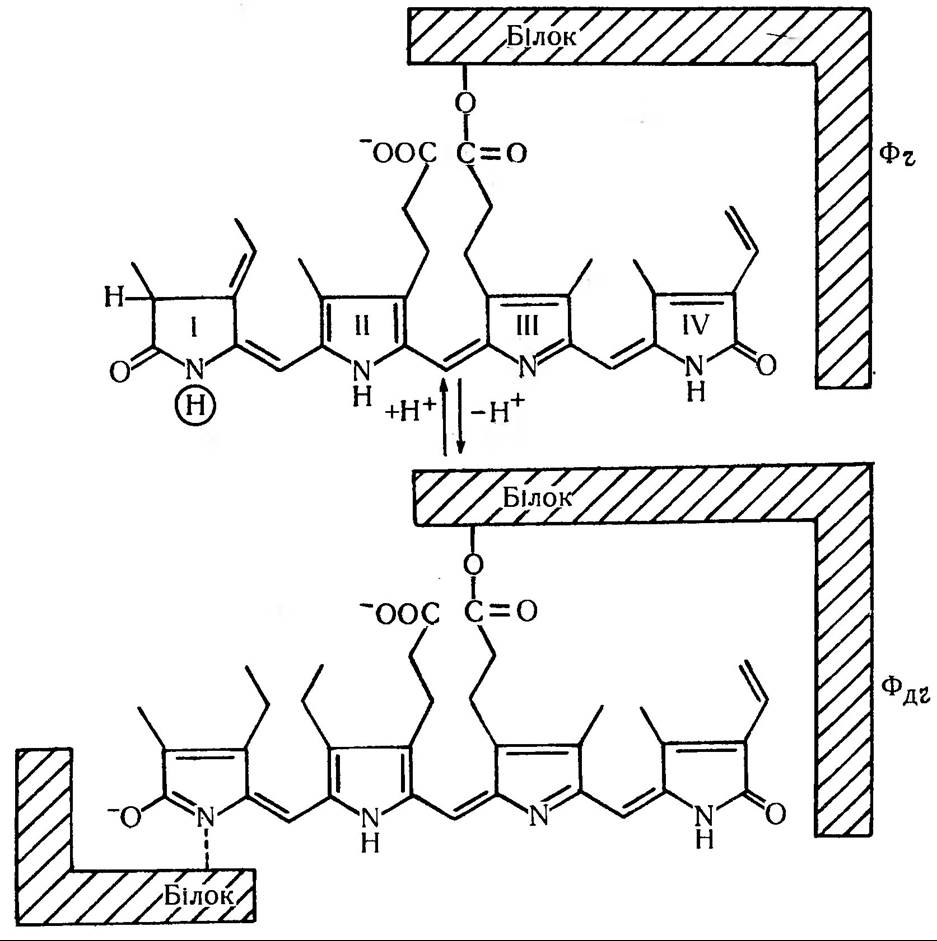
Рис 186. Взаємоперетворення двох форм фітохрому
Сонячне світло містить приблизно однакові долі червоних і далеких червоних променів, тому в рослинному організмі в умовах видимого світла співвідношення форм фітохрому приблизно однакове. Реакції, які керуються фітохромом, залежать від концентрації фітохрому далекого червоного. Звичайно, вони досягають насичення, коли 50% фітохрому перебуває у формі Фд.ч. і тривають до того часу, доки буде його достатня кількість.
Молекула фітохрому складається з двох частин: відносно невеликого хромофору, що поглинає світло, та значно більшого безбарвного білка. Хромофор подібний до фікобілінів і являє собою тетрапірол з незамкнутим ланцюгом. Після поглинання фітохромом світла форма хромофорної групи змінюється залежно від довжини поглинутих променів, а це, в свою чергу, спричинює конформаційні зміни білкового компонента. Білковий компонент складається з 4 субодиниць, молекулярна маса 240 000.
Перетворення Фч→ Фд.ч. при короткочасній дії червоного світла впливає на біохімічні реакції рослин протягом кількох годин наступної темряви. Така разюча стійкість ефекту досить короткого опромінення пояснюється тим, що Фд.ч., будучи активною формою фітохрому, відносно стабільний у темряві. В деяких дослідах активність Фд.ч. в ростучих тканинах спостерігали навіть через 72 год після опромінення червоним світлом. На відміну від фітохрому активовані форми інших пігментів досить нестабільні. Так, молекула збудженого хлорофілу живе, як відомо, кілька мілісекунд.
Повільне зниження рівня Фд.ч. в темряві після опромінення червоним світлом зумовлене двома процесами: реверсією та руйнуванням:

В одних тканинах Фд.ч. в темряві повільно перетворюється на Фч, а в інших він руйнується. В деяких випадках обидва процеси відбуваються одночасно. Процес руйнування фітохрому та його реверсія залежать від температурного фактора (високі температури прискорюють процес). Допускають участь G-протеїнів в активації експресії фітохром-регулюючого гена. Зокрема, виявлено, що плазматичні мембрани етіольованих проростків вівса мають оборотну ГТФ- зв’язуючу активність, залежну від червоних і далеких червоних променів. На цю активність впливає також синє світло.
Як діє фітохром? Фітохрому в рослинній клітині дуже мало, проте незначні кількості поглинутої енергії спричинюють таку різноманітну фізіологічну реакцію. Тому цілком ймовірно, що первинне поглинання фотонів повинно бути сполучене з клітинною реакцією якимось механізмом підсилення. Такий механізм міг би включати управління функцією генів, активністю ферментів, змінювати властивості мембран або вміст таких речовин, як фітогормони, які, в свою чергу, в незначних кількостях впливають на різні фізіологічні процеси. Всі ці процеси вірогідні за певних обставин, але все ж таки основним механізмом підсилення слід вважати регуляцію функцій мембран.
Онтогенетичні реакції, такі як початок цвітіння, проростання насіння та деетіоляція, безперечно, пов’язані з діяльністю рослинної клітини, радикальними змінами в її хімізмі, структурі та функціях. Такі зміни, в свою чергу, залежать від зміни активності ряду ферментів, а також від синтезу їх de novo. У зв’язку з тим, що Ферменти являють собою білки, синтез яких визначається процесами трансляції та транскрипції, стан фітохрому повинен впливати на один з цих процесів або одночасно на обидва. Можливо він зв’язується з ядерним хроматином, впливаючи таким чином безпосередньо на Синтез РНК і білків:
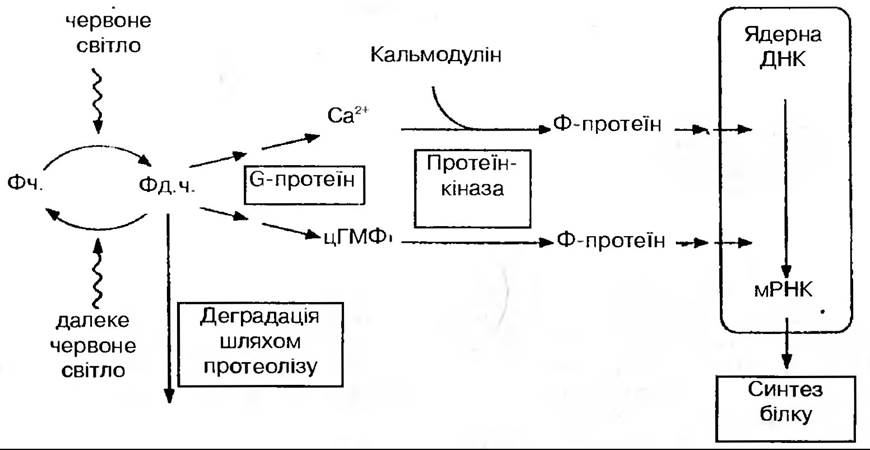
Як уже відмічалось, багато фізіологічних і морфологічних змін, індукованих фітохромом обумовлюється навіть короткочасним освітленням малої ефективності (1/100 сонячного денного освітлення впродовж 1 хв).
Можливо, вплив його ще більш тонкий через зміну компартментації іонів всередині клітини та як результат цього через зміни в синтезі білків.
Однак контроль білкового синтезу — не єдиний спосіб регуляторної дії фітохромів, тому що процеси, які він регулює, не залежать від синтезу білка та відбуваються дуже швидко. Як відомо, рослинні гормони швидко транспортуються по рослині та ефективні в незначних кількостях. Можна припустити, що синтез та розщеплення фітогормонів може бути результатом прямого ферментативного впливу Фд.ч. або впливу його на активність синтетичних та гідролітичних ферментів, які до опромінення існують у вигляді проферментів. Зв’язування чи звільнення фітогормонів в місцях функціонування та зберігання їх може відбуватися в результаті модифікації таких місць фітохромом Фд.ч.
Вміст таких гормонів, як Гібереліни, Цитокініни, Етилен, ауксин, в тканинах рослин, вирощених у темряві, швидко змінюється після короткого опромінення червоним світлом, тоді як рівень абсцизової кислоти змінюється при довготривалому освітленні червоним світлом. У зв’язку з тим що вплив червоного світла на гібереліни, цитокініни та етилен можна зняти далеким червоним світлом, фоторецептором для цих ефектів, безперечно, служить фітохром. Фд.ч. підвищує, як відомо, рівень гіберелінів та цитокінінів і, навпаки, знижує вміст ауксину та етилену.
Швидка ефективність його дії, мабуть, пояснюється саме впливом його на структуру та функції мембран. Гіпотеза про те, що фітохром діє на рівні мембран, підкріплюється також прямим демонструванням включення фітохромів та функціонуванням його в штучних ліпідних мембранах. Опромінення таких мембран червоним та далеким червоним світлом спричинює значні зміни в їхніх фізичних властивостях (наприклад, електричний опір). Це свідчить про те, що кон- формаційні зміни хромофора фітохрому можуть одночасно змінювати і структуру мембрани. Інші, більш віддалені ефекти (або активація генів) можуть також перебувати під впливом попередніх проявів дії фітохрому.
Таким чином, виходячи з того, що фітохром повязаний з мембраною, можна виділити декілька можливих механізмів його дії:
·регуляція активного транспорту іонів і молекул через мембрану (можливо через зміну активності АТФ-ази);
·регуляція активності фітогормонів, зв’язаних з мембраною;
·вплив на активність зв’язаних з мембраною білків.
Слід пам’ятати, що не всі реакції, що контролюються фітохромом, локалізовані в межах тієї самої ж клітини. В деяких випадках опромінення однієї частини рослини впливає на розвиток органів, які знаходяться на певній відстані. Наприклад, фотоперіодична індукція цвітіння, формування бульб, перехід до стану спокою. Тому будь-яка гіпотеза про механізм дії фітохрому повинна пояснювати не лише локальні, а й просторово віддалені реакції.
Нарешті, деякі фотоморфогенетичні реакції потребують значно більше світлової енергії і спричинюються переважно синім або далеким червоним світлом. Наприклад, перехід до цвітіння, утворення антоціанів може регулюватися фітохромом, або ще якимось пігментом, що поглинає сині промені. Можливо, синє світло поглинає якийсь інший пігмент, наприклад флавопротеїд.
В останній час висловлюються думки, що фітохром може виступати і як детектор, що уловлює затінення іншими рослинами. Випромінювання 700 нм майже повністю відбивається або поглинається рослиною, тоді як далеке червоне світло (700-800 нм) значною мірою пропускається. Це спричинює критичний зсув вгору відношення Фч до Фд.ч., тобто переходить більше Фд.ч. у затінених рослин, що зумовлює швидке збільшення росту міжвузлів.
Реакції на червоне (далеке червоне світло) мають відношення і до синтезу антоціанів у яблук, капусти та до великого різноманіття інших реакцій рослин на всіх фазах життєвого циклу. Щодо такого складного явища, як Фотоперіодизм, то воно поряд із взаємоперетворенням різних форм фітохрому контролюється й іншими факторами.
Механізм дії синього світла. Фітохром має незначний пік поглинання у фіолетовій і синій області спектра (460-475 нм). Відомо, що додавання цих променів до червоного світла посилює синтез амінокислот і білків у рослин, причому дія синього світла має регуляторний характер. Однак у більшості випадків фізіологічна дія синього світла не залежитиь від далекого червоного світла. В таких випадках синє світло діє не через систему фітохрому, а через криптохром. Синє світло часто викликає гіперполяризацію мембранного потенціалу клітин, впливає на функціонування редокс-ланцюгів, локалізованих у внутрішньоклітинних мембранах (ендоплазматичний ретикулум, Апарат гольджі, Мітохондрії) та плазмалемі клітин. Флавінові компоненти (флавопротеїни) редокс-ланцюгів забезпечують поглинання синього світла, тим самим впливають на метаболізм, перш за все контролюючи іонні потоки. Останні, в свою чергу, є учасниками механізмів регуляції росту і морфогенезу.
Таким чином, кінцева форма зеленої рослини, що зумовлена в цілому його генами, визначається за участю світла, яке діє через різні фоторецептори. Бо протохлорофіл регулює процес зеленіння, хлорофіл — загальний автотрофний тип живлення, фітохром, а також якийсь інший пігмент типу флавопротеїдів — різноманітні тонкі внутрішньоклітинні процеси. Зелена рослина — воістину машина, що приводиться в дію світлом і живе в постійній залежності від падаючих на неї квантів.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.