Медична генетика - В. М. Запорожан 2005
Етіологія спадкових хвороб
Типи мутацій, обумовлених зміною кількості й структури хромосом
Хромосомні аберації
Всі Мутації, які пов’язані зі зміною кількості й структури хромосом, можна розділити на три групи:
— хромосомні аберації, зумовлені зміною структури хромосом;
— геномні мутації, зумовлені зміною кількості хромосом;
— міксоплоїдії — мутації, зумовлені наявністю різних за хромосомними наборами клонів клітин.
Хромосомні аберації (хромосомні мутації) — це зміни в структурі хромосом (схема 2.3), які є, як правило, наслідком нерівного кросинговеру при мейозі. До хромосомних аберацій призводять також розриви хромосом, спричинені іонізуючою радіацією, деякими хімічними мутагенами, вірусами та іншими мутагенними факторами. Хромосомна аберація може бути незбалансованою і збалансованою.
При незбалансованих мутаціях відбувається втрата або збільшення генетичного матеріалу, змінюється кількість генів або їх активність, що спричинює зміну фенотипу.
Хромосомні перебудови, які не спричинюють зміни генів або їхньої активності і не змінюють фенотип, називаються збалансованими. Проте хромосомна аберація порушує кон’югацію хромосом і Кросинговер при мейозі, що викликає появу гамет із незбалансованими хромосомними мутаціями. У носіїв збалансованих хромосомних аберацій може бути Безплідність, висока частота спонтанних абортів, високий ризик народження дітей із хромосомними хворобами.
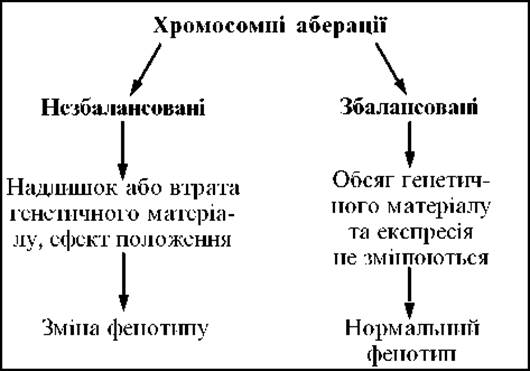
Схема 2.3. Фенотипічний ефект хромосомних аберацій
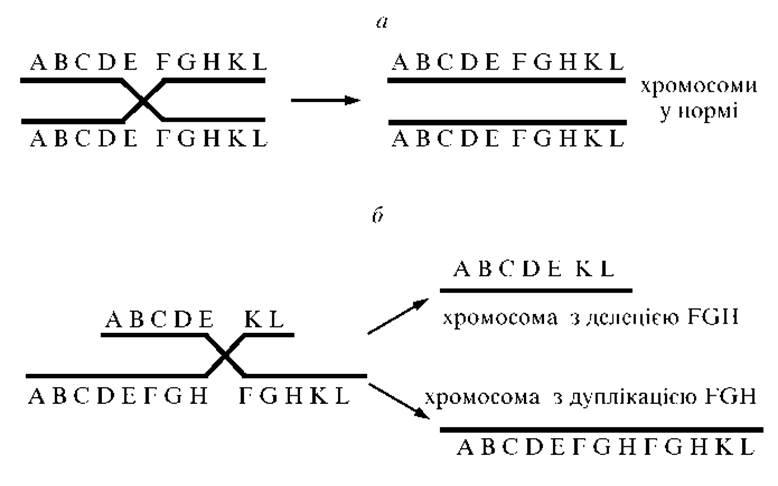
Рис. 2.2. Механізм формування хромосом з делецією і дуплікацією внаслідок нерівного кросинговеру:
а — схема кросинговеру в нормі; б — Нерівний кросинговер — утворення хромосом з делецією і дуплікацією
Виділяють такі типи хромосомних мутацій:
1. Делеція, або брак — втрата ділянки Хромосоми.
![]()
2. Дуплікація — подвоєння ділянки хромосоми.
![]()
Делеції і дуплікації часто є наслідком порушення процесу кросинговеру (рис. 2.2).
3. Інверсія — поворот ділянки хромосоми на 180° (в одній з ділянок хромосоми гени розташовані в послідовності, зворотній порівняно з нормальною). Якщо інвертована ділянка включає центромеру, то інверсія називається перицентричною, якщо не захоплює — парацентричною (рис. 2.3).
Якщо в результаті інверсії не змінюється кількість хромосомного матеріалу і немає ефекту положення, то індивіди фенотипічно здорові. Часто зустрічається перицентрична інверсія 9-ї хромосоми, яка не приводить до зміни фенотипу. При інших інверсіях можуть порушуватися кон’югація і кросинговер, внаслідок чого відбувається розрив хромосом і утворення незбалансованих гамет.
4. Кільцева хромосома (рис. 2.4) виникає при втраті двох теломерних фрагментів. «Липкі» кінці хромосоми з’єднуються, утворюючи кільце (рис. 2.5).
Ця мутація може бути як збалансованою, так і незбалансованою (залежно від об’єму хромосомного матеріалу, який втрачається).
Кільцеві хромосоми нестабільні, оскільки при редуплікації виникають подвійні кільця, які потім розриваються. У гаметах відбуваються різні хромосомні перебудови (рис. 2.6).
5. Ізохромосоми (рис. 2.7) — втрата одного плеча хромосоми і дуплікація другого. Найімовірніше вони виникають внаслідок горизонтального, а не поздовжнього поділу центромери.
В результаті утворюється метацентрична хромосома, що має два однакових плеча. Частіше зустрічається ізохромосома за довгим плечем Х-хромосоми. Каріотип записують: 46,Х,i(Xq). Ізохромосома Х спостерігається в 15 % усіх випадків синдрому Шерешевського — Тернера.
6. Транслокація — перенесення ділянки хромосоми на негомологічну хромосому, в іншу групу зчеплення. Виділяють кілька типів транслокацій:
а) реципрокні транслокації — взаємний обмін ділянками між двома негомологічними хромосомами (рис. 2.8, а).
У популяціях частота реципрокних транслокацій 1:500. З нез’ясованих причин частіше зустрічається реципрокна транслокація, що залучає довгі плечі 11-ї та 22-ї хромосом. У носіїв збалансованих реципрокних транслокацій часто бувають спонтанні аборти або народжуються діти з множинними вродженими вадами розвитку внаслідок утворення гамет з незбалансованими мутаціями. Генетичний ризик у носіїв таких транслокацій коливається від 1 до 10 %;
б) нереципрокні транслокації (транспозиції) — переміщення ділянки хромосоми або всередині тієї ж хромосоми (рис. 2.8, б), або в іншу хромосому без взаємного обміну;
в) особливий вид транслокацій — робертсонівська транслокація (або центричне злиття).
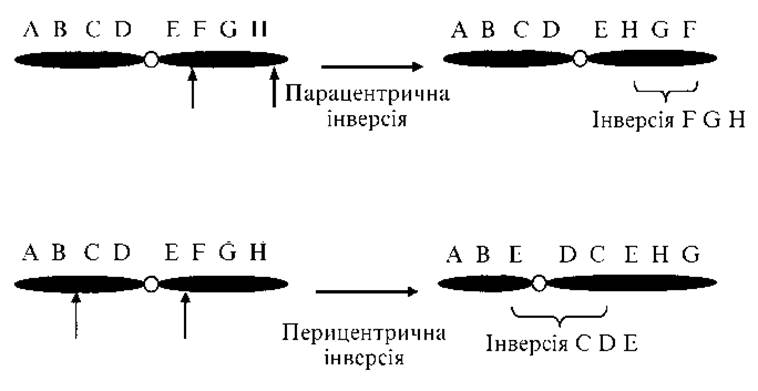
Рис. 2.3. Парацентрична і перицентрична інверсії
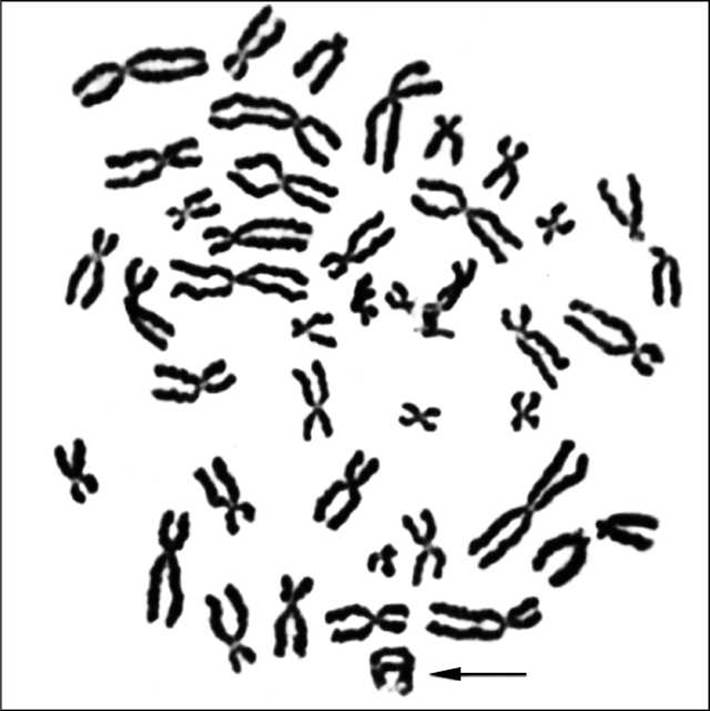
Рис. 2.4. Кільцева хромосома у метафазній пластинці (відмічена стрілкою)
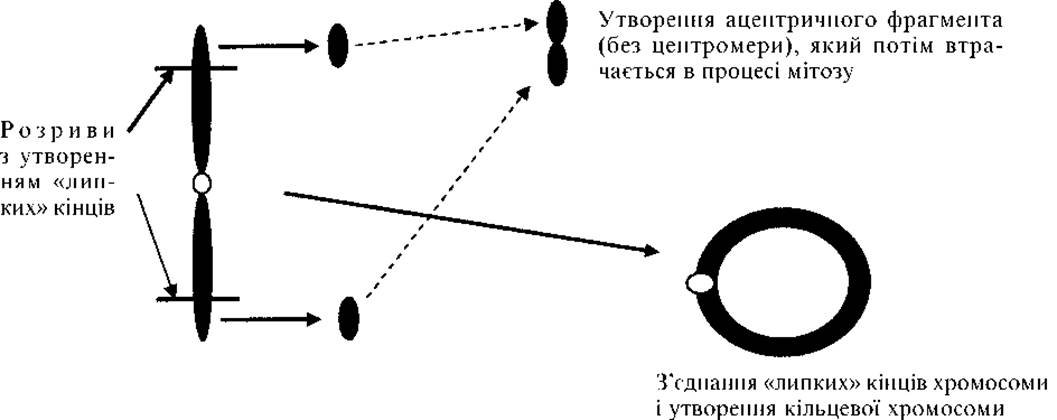
Рис. 2.5. Механізм формування кільцевої хромосоми
Спостерігаються між будь-якими двома акроцентричними хромосомами з групи D (13, 14 і 15-та пари) і G (21-ша і 22-га пари). При центричному злитті дві гомологічні або негомологічні хромосоми втрачають короткі плечі й одну центромеру, довгі плечі з’єднуються (рис. 2.9). Замість двох хромосом утворюється одна, що містить генетичний матеріал довгих плечей двох хромосом. Загальна кількість хромосом у носіїв збалансованої робертсонівської транслокації — 45. У коротких плечах всіх десяти хромосом груп D і G знаходяться однакові гени, що кодують рРНК. Кожна Клітина має велику кількість (до 105) копій цих генів. Втрата коротких плечей двох хромосом не приводить до істотних змін фенотипу, оскільки втрата генів компенсується роботою таких же генів решти восьми акроцентричних хромосом.
Таким чином, носії робертсонівських транслокацій здорові, але у них підвищена частота спонтанних абортів і високий ризик народження дітей із хромосомними хворобами. Частота робертсонівських транслокацій у популяції становить 1:1000.
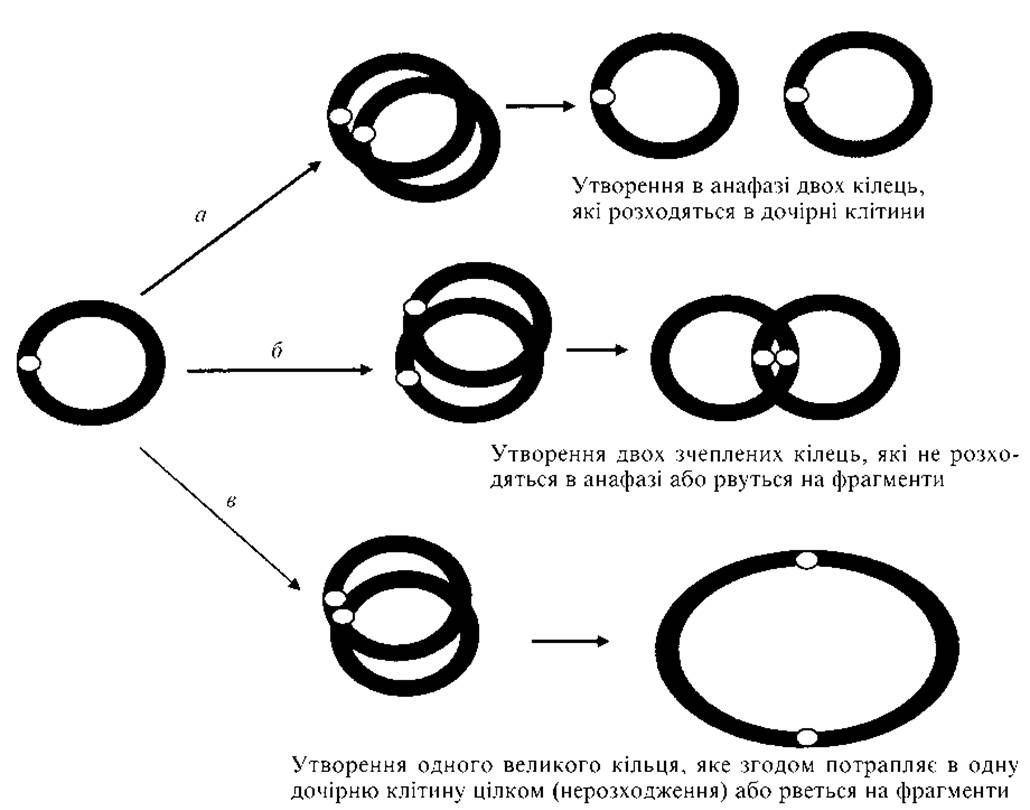
Рис. 2.6. Варіанти розходження кільцевої хромосоми в мітозі:
а — нормальна реплікація; б — реплікація і два сестринських обміни при кросинговері; в — редуплікація і один сестринський обмін при кросинговері
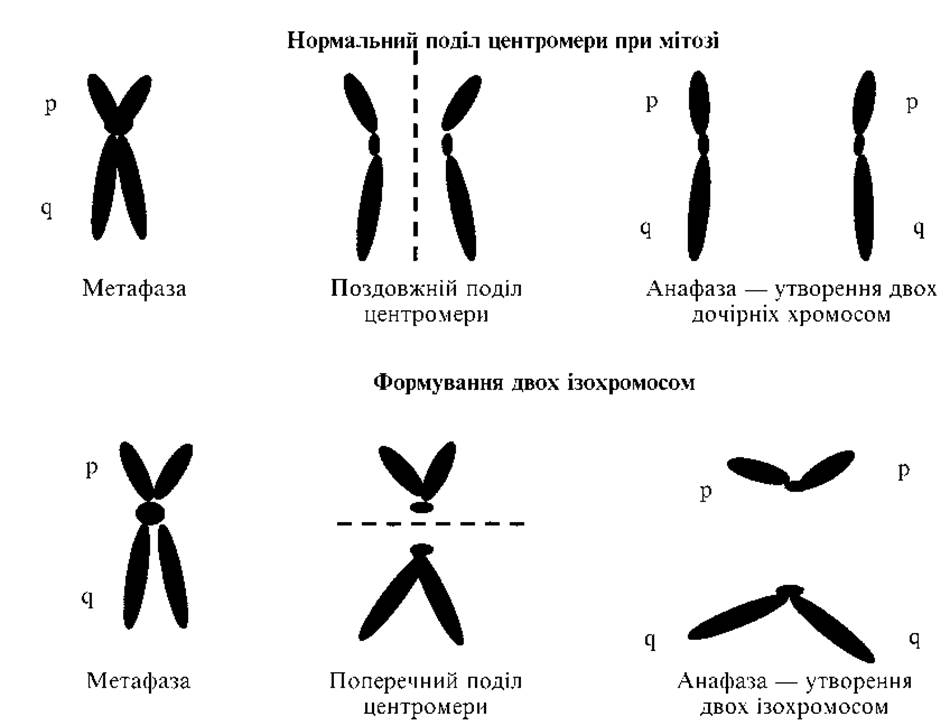
Рис. 2.7. Механізм формування ізохромосоми
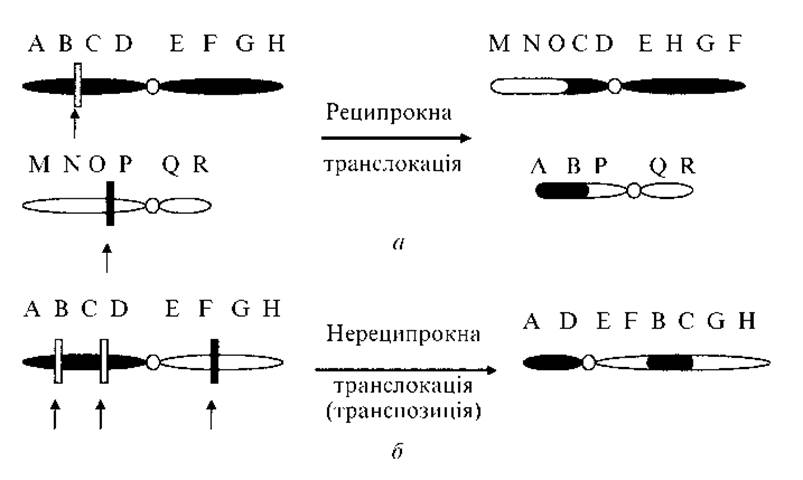
Рис. 2.8. Механізм формування транслокацій:
а — реципрокна транслокація; б — нереципрокна транслокація
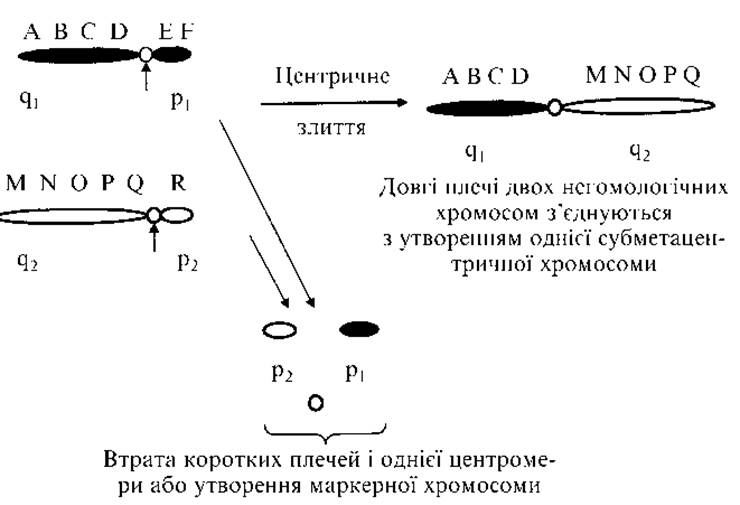
Рис. 2.9. Центричне злиття хромосом (робертсонівська транслокація)
Прикладом може бути злиття довгих плечей 14-ї та 21-ї хромосом (14q21q). Носій такої збалансованої транслокації має тільки 45 хромосом і нормальний фенотип (рис. 2.10). Людина з нормальним каріотипом має дві хромосоми з 14-ї пари і дві — з 21-ї (14,14,21,21). Гамети в нормі мають одну хромосому 14 і одну 21 (14,21). У носія збалансованої транслокації замість чотирьох хромосом буде три (14,14q21q,21).
Носії теоретично утворюють 6 типів гамет (табл. 2.3) з неоднаковою вірогідністю утворення.
Іноді один із батьків є носієм збалансованої транслокації, при якій спостерігається центричне злиття двох гомологічних хромосом групи D або G. У таких людей утворюється два типи гамет. Наприклад, при транслокації 21q21q утворюються гамети:
1) 21q21q;
2) 0 — тобто гамета без хромосоми 21.
Після Запліднення нормальною гаметою утворюється два типи зигот: 1) 21,21 q21 q — транслокаційна форма синдрому Дауна; 2) 21,0 — моносомія 21-ї хромосоми, летальна мутація. Вірогідність народження хворої дитини становить 100 %.
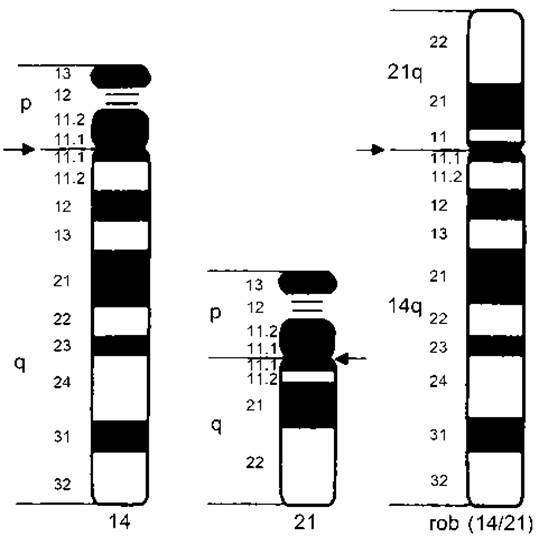
Рис. 2.10. Збалансована робертсонівська транслокація хромосом 14 і 21 (носій має 3 хромосоми: нормальну 14-ту, нормальну 21-шу і хромосому, яка об’єднує довгі плечі 14-ї і 21-ї хромосом)
Таблиця 2.3. Можливі типи гамет і зигот у носія збалансованої транслокації 14q21q
|
Можливі типи гамет у носія збалансованої транслокації з каріотипом 14,14q21q,21 |
Каріотип зигот, які утворюються після злиття можливих типів гамет з клітиною, що містить нормальний набір хромосом (14,21) |
|
14,21 — нормальний набір хромосом |
14,14,21,21 — нормальний каріотип |
|
14q21q — збалансована хромосомна мутація |
14,14q21q,21 — збалансована хромосомна мутація, нормальний фенотип |
|
14,14q21q — гамета з незбалансованою мутацією (зайве довге плече 14-ї хромосоми) |
14,14,14q21q,21 — незбалансована хромосомна мутація із зайвим довгим плечем 14-ї хромосоми, летальна мутація, яка призводить до загибелі ембріона на ранніх етапах ембріонального розвитку |
|
14q21q,21 — гамета з незбалансованою мутацією (зайве довге плече 21-ї хромосоми) |
14,14q21q,21,21 — транслокаційна форма синдрому Дауна |
|
14 — відсутня хромосома 21-ї пари |
14,14,21 — моносомія по 21-й хромосомі, летальна мутація для статевих клітин або ранніх ембріонів |
|
21 — відсутня хромосома 14-ї пари |
14,21,21 — моносомія по 14-й хромосомі, летальна мутація для статевих клітин або ранніх ембріонів |

Делеції та дуплікації змінюють кількість генів у організмі (часткова моносомія або трисомія). Інверсії, транслокації, транспозиції змінюють розташування генів у хромосомах.
7. Центричне розділення — явище, зворотне центричному злиттю. Одна хромосома ділиться на дві, при цьому повинна утворитися нова центромера. Інакше хромосома без центромери втрачається при поділі клітин.
8. Дицентричні хромосоми (рис. 2.11) містять дві центромери. Дві дочірні хроматиди втрачають теломери й об’єднуються в одну хромосому (рис. 2.12). При мітозі та мейозі порушується розходження таких хромосом, виникають розриви.
9. Маркерна хромосома — це додаткова хромосома (вірніше, фрагмент якої-небудь хромосоми з центромерою). Зазвичай має вигляд дуже короткої акроцентричної хромосоми, рідше — кільцевої або іншої. Якщо маркерна хромосома містить тільки гетерохроматин, то фенотип не змінюється. Якщо ж вона містить еухроматин (експресовані гени), то це поєднано з розвитком хромосомної хвороби (аналогічно дуплікації якої-небудь ділянки хромосоми).

Рис. 2.11. Дицентрична хромосома в метафазній пластинці (відмічена стрілкою)
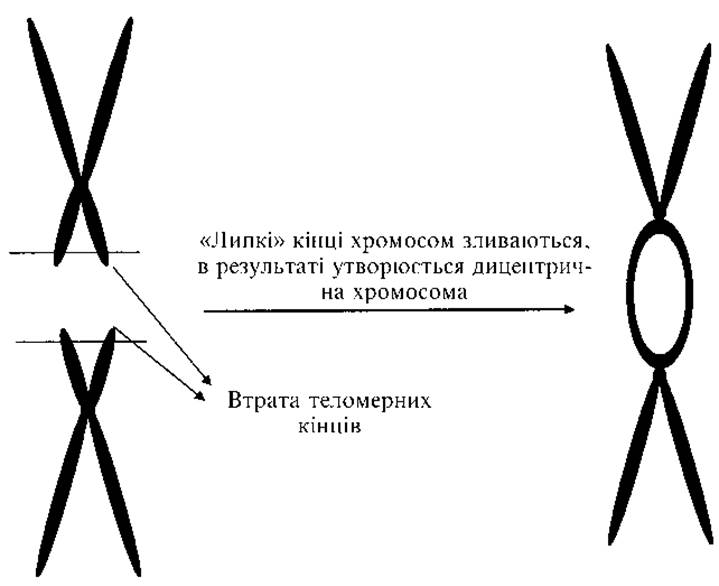
Рис. 2.12. Механізм формування дицентричної хромосоми
Значення хромосомних мутацій в еволюції
Хромосомні мутації відіграють важливу роль в еволюції. В процесі еволюції відбувається активна перебудова хромосомного набору за допомогою інверсій, робертсонівських транслокацій та інших мутацій. Чим далі один від одного відстоять організми, тим більше відрізняються їхні хромосомні набори.
Наприклад, у процесі формування людини від мавп відбулася принаймні одна робертсонівська перебудова. У людини 23 пари хромосом, а у великих людиноподібних мавп — 24. Два плеча великої другої Хромосоми людини відповідають двом різним хромосомам мавп (це хромосоми 13 і 14 горили та орангутанга). Хромосоми 4, 5, 12 і 17 людини і шимпанзе відрізняються між собою перицентричними інверсіями.
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.