ЦИТОЛОГІЯ, ГІСТОЛОГІЯ, ЕМБРІОЛОГІЯ - 2008
Розділ 3. ЗАГАЛЬНА ГІСТОЛОГІЯ
3.4 М'язова тканина
3.4.3 Серцева м'язова тканина (міокард)
Серцева М'язова тканина складається із м'язових клітин-кардіоміотитів, які, з'єднуючись своїми кінцями по довгій осі клітини формують структури, подібні до м'язових волокон. Поперечна смугастість має ту ж природу, що і в скелетних м'язах, тобто зумовлена оптичною неоднорідністю міофібрил, що складаються із двох типів міофіламентів. Останні містять анізотропні A-диски та ізотропні І-диски, що займають периферійну частину клітини. Кардіоміоцити формують м'язові волокна, які анастомозують між собою, утворюючи симпласт.
Різниця в будові та функції кардіоміоцитів дає підставу класифікувати їх на два види: Скоротливі, або типові серцеві міоцити (становлять більшу частину серцевого м'яза) — це робоча мускулатура та провідні або атипові серцеві міоцити, що формують провідну систему серця.
Типові кардіоміцити — це одно або двоядерні клітини, світлі ядра яких локалізуються у центрі (рис.66), на відміну від крайового розміщення у скелетних м'язових волокнах. На поверхні міоцити мають відростки, або анастомози, за допомогою яких вони з'єднуються. Саркоплазма містить міофібрили, Органели, включення, гіалоплазму. Добре розвинені Мітохондрії (саркосоми), гірше-комплекс Гольджі та сарко- плазматична сітка, яка не утворює великих термінальних цистерн (як у скелетних м'язах). Із включень найбільше міститься гранул глікогену та пігменту ліпофусцину, кількість якого з віком збільшується.
. 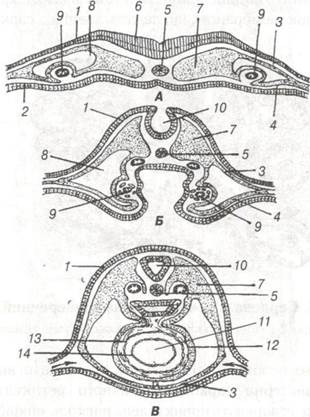
Рис. 65. Гістогенез серця:
А, Б, В — поперечні розрізи зародків на трьох стадійних формуваннях трубчастої закладки серця;
А — дві парні закладки серця; Б — зближення закладок; В — злиття в одну непарну закладку серця:
1-ектодерма; 2-ентодерма; 3- паріетальний листок мезодерми; 4-вісцеральна мезодерма; 5-хорда; 6-нервова пластинка;
7-соміти; 8-вторинна порожнина тіла (целом); 9-ендотеліальна закладка; 10-нервова трубка; 11-порожнина серця;
12- епікард; 13 — міокард; 14-ендокард
Місце з'єднання міоцитів - це вставні ducки-aналоги Z-плacтuнок (телофрагми). Вставні диски (рис. 67) під електронним мікроскопом мають хвилястий вигляд і два види сполучення: це десмосоподібні та щільні контакти. Десмосоподібні контакти забезпечують міцне з’єднання клітин, а щільні-електричний зв’язок сусідніх кардіоміоцитів та проведення імпульсів по м'язовому пласту міокарда. У кардіоміоцитах Т-система (трубочки) заходять всередину клітини на рівні Z-пластинок (телофрагм), тому їх кількість відповідає кількості саркомерів. Т-трубочки набагато ширші, ніж у скелетних м'язах і, крім того, ще вистелені базальною мембраною, що лежить зовні від сарколеми.
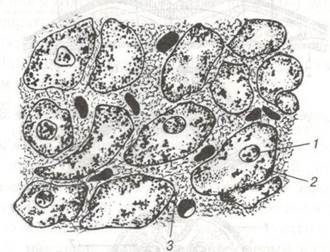
Рис. 66. Серцева м'язова тканина (поперечний зріз);
1-Ядро; 2-Цитоплазма клітини; 3-Пухка сполучна тканина.
Характерною особливістю серцевих міоцитів є відсутність тріад, оскільки цистерни саркоплазматичного ретикулуму разом із Т-трубочками не утворюють нових кілець навколо міофібрил. Функція Т-трубочок кардіоміоцитів - це проведення імпульсів у клітину і забезпечення одночасного скорочення усіх міофібрил. Оскільки у типових серцевих міоцитів відсутні тріади, для їх скорочення необхідний приплив іонів кальцію з інших тканин. Ось чому виведення в Кров солей кальцію підсилює роботу серцевого м'яза. Кардіоміоцити, що розміщуються у передсерді, мають здатність синтезувати передсердний натрійуретичний фактор (ПНФ), який діє діуретично (підсилює виведення з організму води і солей, та знижує артеріальний тиск). Передсердний натрійуретичний фактор — це поліпептидний гормон, що є модулятором або антагоністом системи ренін-ангіотензин-альдостерон (гормони нирок і надниркових залоз).
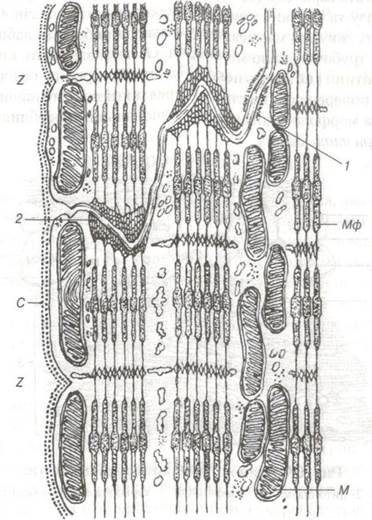
Рис. 67 Схема ультраструктури серцевого м'яза в ділянці вставної смужки:
С — сарколема; М — мітохондрії; Мф — міофіламенти:
1-зона ущільнення на плазмолемі; 2-закінчення міофіламентів на плазмолемі; Z — вставна смужка.
Атипові, провідні кардіоміоцити формують провідну систему серця (рис. 68). У різних видів тварин вони мають різну будову й, особливо, у тварин із сповільненим пульсом. Ці клітини грушоподібної, видовженої форми, з великою кількістю анастомозів. Ядра містять незначну кількість гетерохроматину, добре виражене ядерце, локалізуються у центрі клітини. Саркоплазма містить велику кількість глікогену і небагато саркосом (мітохондрій), свідчить про інтенсивний процес гліколізу та низький рівень окислювальних процесів. Саркоплазматичний ретикулум має незначну кількість рибосом, слаборозвинену систему Т-трубочок, мало міофібрил. Останні займають периферійну частину клітини і не мають певної орієнтації, через що недостатньо виявляється поперечна смугастість. Серед провідних, атипових серцевих міоцитів за морфологічними та функціональними особливостями виділяють три типи клітин.
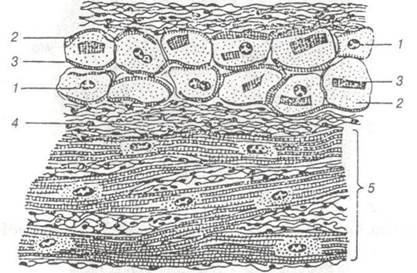
Рис. 68. Атипові, провідні кардіоміоцити:
1- ядро; 2- цитоплазма; 3 — міофібрили; 4- саркоплазма; 5- робочі кардіоміоцити.
Перший тип - пейсмекерні клітини (Р-клітини) — це водії ритму серця. Вони мають нестабільний біопотенціал спокою і здатні у спокої деполяризуватися з частотою 70 разів за 1 хв, тобто ці клітини генерують імпульси до скорочення. Вони знаходяться в центральній частині синусно-передсердного вузла, мають багатокутну форму, діаметр 8-10 мкм. і невелику кількість міофібрил. Саркоплазматична сітка розвинена слабо, Z-система відсутня, є багато піноцитозних пухирців та кавеол.
Клітини другого типу - це перехідні клітини, котрі передають збудження від P-клітин до пучка скоротливих елементів міокарду. Вони локалізуються на периферії синусно-передсердного вузла і утворюють більшу частину передсердно-шлункового вузла. Ці клітини мають витягнуту форму з більшою кількістю міофібрил ніж у Р-клітин.
Клітини третього типу - це клітини пучка провідної системи (волокон Пуркіне) вони передають збудження від перехідних клітин до скоротливих міоцитів шлуночків. Мають діаметр до 15 мкм., невелику кількість міофібрил, значну кількість глікогену та ензими анаеробного гліколізу.
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.