Фізіологія рослин - Мусієнко М.М. 2001
Фотосинтез: фізіолого-біохімічні та екологічні аспекти
Листок як орган фотосинтезу
Особливості морфології, анатомії та фізіології листка повною мірою забезпечують здійснення його основної функції — фотосинтезу (рис. 33). Листки, в більшості випадків тонкі
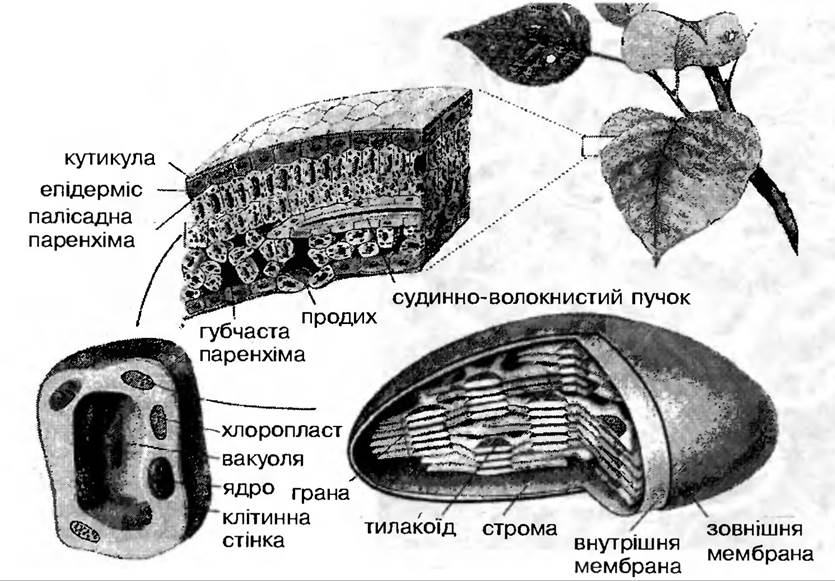
Рис. 33. Листок як орган фотосинтезу
і володіють максимальною поверхнею на одиницю маси. Загальна листкова поверхня рослин помірної зони, розміщених на 1 га орної площі, майже в 5 разів перевищує земельну ділянку, а в південних районах з нормальним зволоженням — в 10-15 разів.
Різноманітна форма їх, розташування на стеблі, фототропічна реакція забезпечують виконання найважливішої функції — поглинання світлових променів. Як і всі фізичні тіла, листок відбиває, поглинає та пропускає падаючі на нього промені (рис. 34).
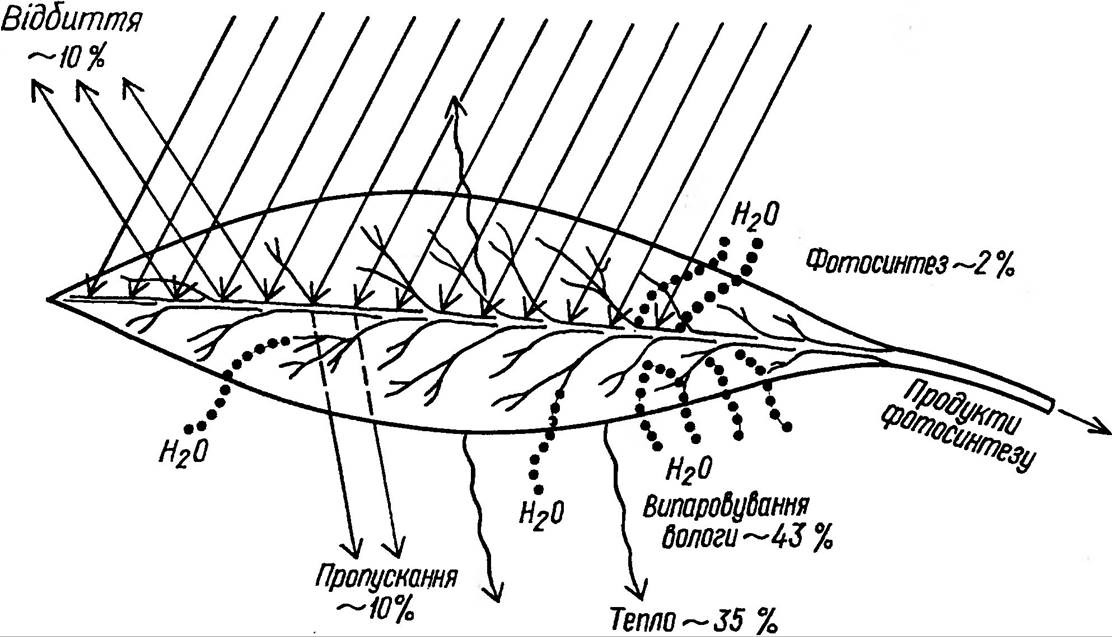
Рис. 34. Енергетичний баланс листка
Поверхня листка вкрита епідермісом. Клітини нижнього та верхнього епідермісу, за винятком тих, що створюють продиховий апарат, позбавлені хлоропластів. Вони лише пропускають світлові промені, отже, безпосередньої участі в фотосинтезі не беруть.
Характерною особливістю покривної Тканини листка є наявність продихів, площа яких не перевищує 1% загальної поверхні листка. Замикаючі клітини продихів мають тонкі зовнішні та товсті внутрішні клітинні стінки. При зміні тургору такі стінки нерівномірно розтягуються, що сприяє відкриванню їх чи, навпаки, закриванню (рис. 35). Для того щоб процес фотосинтезу відбувався нормально, в клітини до зелених пластид повинен надходити вуглекислий газ. Кількість його в атмосферному повітрі складає близько 0,03%. Впродовж дня рослина засвоює кількість СО2, яка міститься приблизно в 30-60 метровому шарі повітря. Для утворення 1 г вуг-
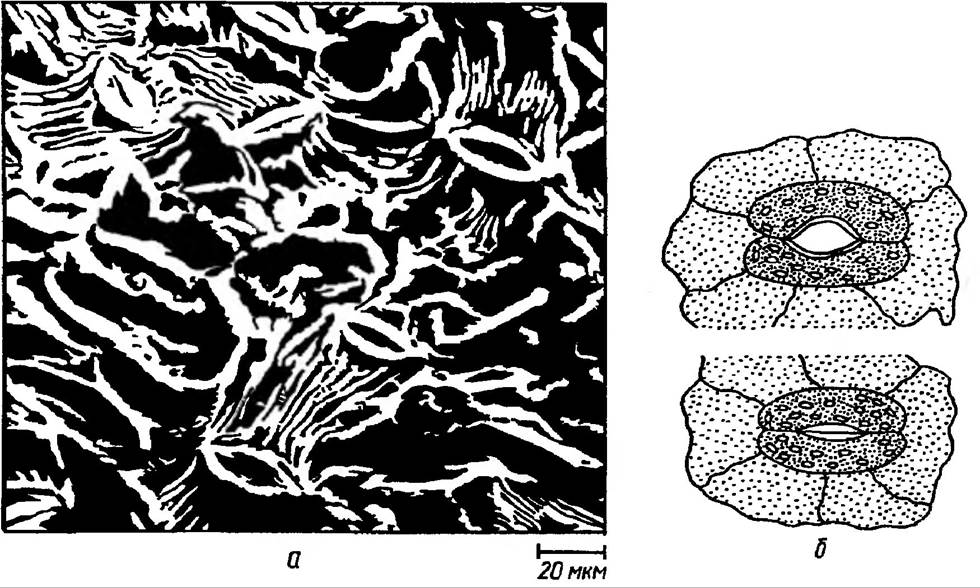
Рис. 35. Продиховий апарат листка:
а — мікрофотографія, одержана за допомогою скануючого електронного мікроскопа, б — схема відкритого та закритого продиху
леводів необхідно близько 1,47 г СО2 (така кількість його міститься майже у 2500 л повітря). Це можливо завдяки безперервному турбулентному рухові повітряних мас навколо листків, який спричинюється нерівномірним нагріванням їх сонячними променями та вітром.
Кількість води, яка витрачається на Фотосинтез, становить зовсім незначну частину тієї кількості, яку поглинає та випаровує рослинний Організм. Дефіцит води знижує швидкість фотосинтезу, адже закриття продихів припиняє доступ вуглекислого газу до мезофілу листка.
Крім того, кисень, що виділяється в процесі фотосинтетичних реакцій фотолізу води, теж потрапляє в навколишнє середовище крізь продихи. Тому закривання продихів припиняє газообмін з атмосферою, хоча при цьому ні фотосинтез, ні Дихання не припиняються. Будучи замкненими в закритому циклі всередині листка, вони взаємно доповнюють один одного, поста- чаючи кисень та вуглекислий газ для відповідних реакцій.
Отже, функціонування продихів регулює надходження вуглекислого газу, виділення кисню та випаровування води. Як правило, продихів більше з нижнього боку листка, куди не потрапляють прямі сонячні промені. На верхній частині їх значно менше, що зменшує випаровування води під дією тих самих променів. Основну ж тканину листка займає мезофіл, який буває двох типів: губчастий та палісадний (рис. 36).
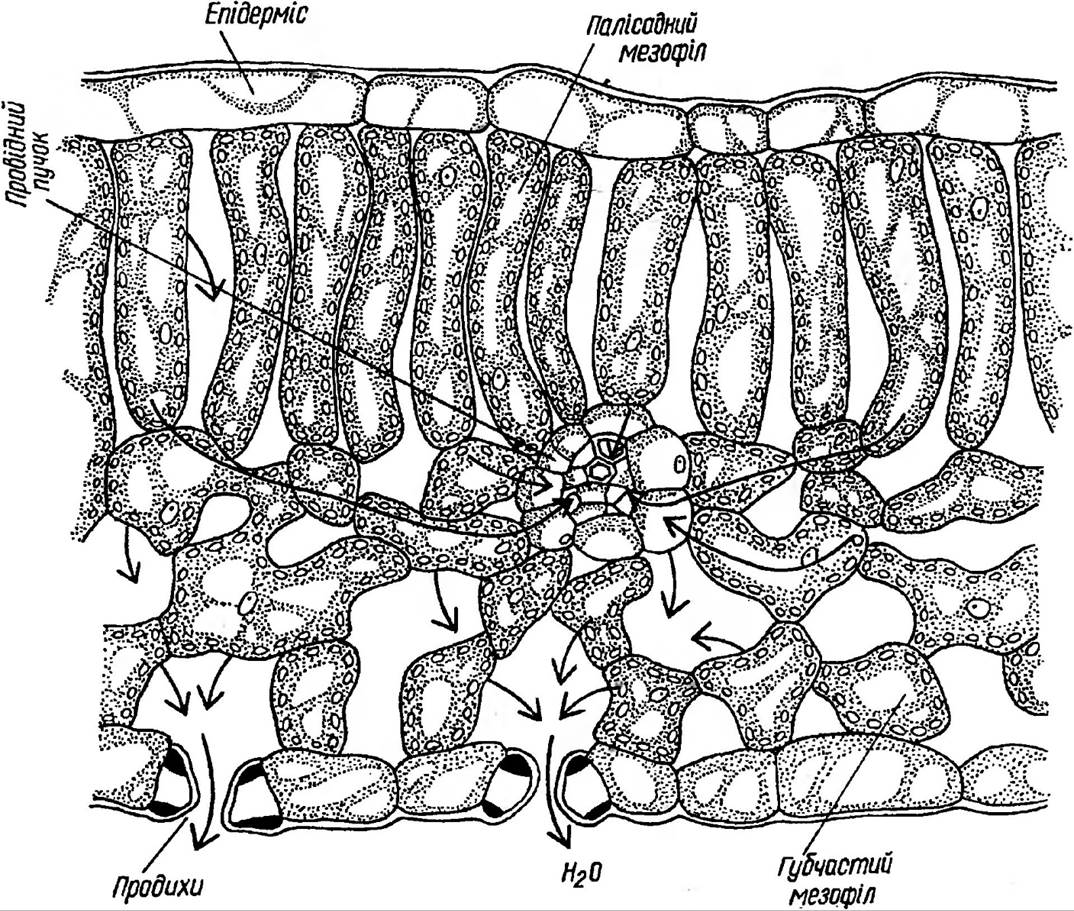
Рис. 36. Анатомічна структура листка
Найбільше хлоропластів мають клітини палісадної або, як її ще називають, стовбчастої паренхіми (кілька десятків на 1 клітину). Ці клітини щільно прилягають одна до одної, на відміну від пухко розміщених клітин губчастого мезофілу. Від 15 до 20% загального об’єму листка становить міжклітинний простір, наявність якого характерна насамперед для губчастих тканин. Завдяки міжклітинникам значно зростає внутрішня поверхня листка (в 7-10 разів вона більша за зовнішню поверхню листка), створюється відповідне співвідношення газового складу, яке відрізняється від атмосферного, хоча вони й поєднуються через продихи. Саме за рахунок міжклітинного простору здійснюється постійне надходження СО2 з атмосфери, бо губчаста тканина мезофілу розміщується поблизу нижнього епідермісу, де знаходяться численні продихи.
Мезофіл листка пронизаний густою сіткою судинно-волокнистих пучків, до складу яких входить ксилема, яка постачає воду та мінеральні речовини, і флоема, по якій відбувається відтік продуктів фотосинтезу.
Характерно, що прямого контакту між ними та кожною клітиною немає. Це свідчить про те, що внутрішнє транспортування всього необхідного для фотосинтезу, як і відтік його продуктів на короткі відстані, здійснюється також по симпласту та апопласту.
Таким чином, щоб фотосинтез відбувався в оптимальному режимі, листок повинен одержувати достатню кількість світлової енергії, води та вуглекислого газу.
Хлоропласти. Фотосинтез здійснюється при безперервному надходженні світлової енергії в значній кількості в строго певні ділянки клітин. Тому в процесі еволюції рослин в клітині виникли спеціалізовані структури — хлоропласти (рис. 37). Розмір хлоропластів коливається від 4 до 10 мкм.
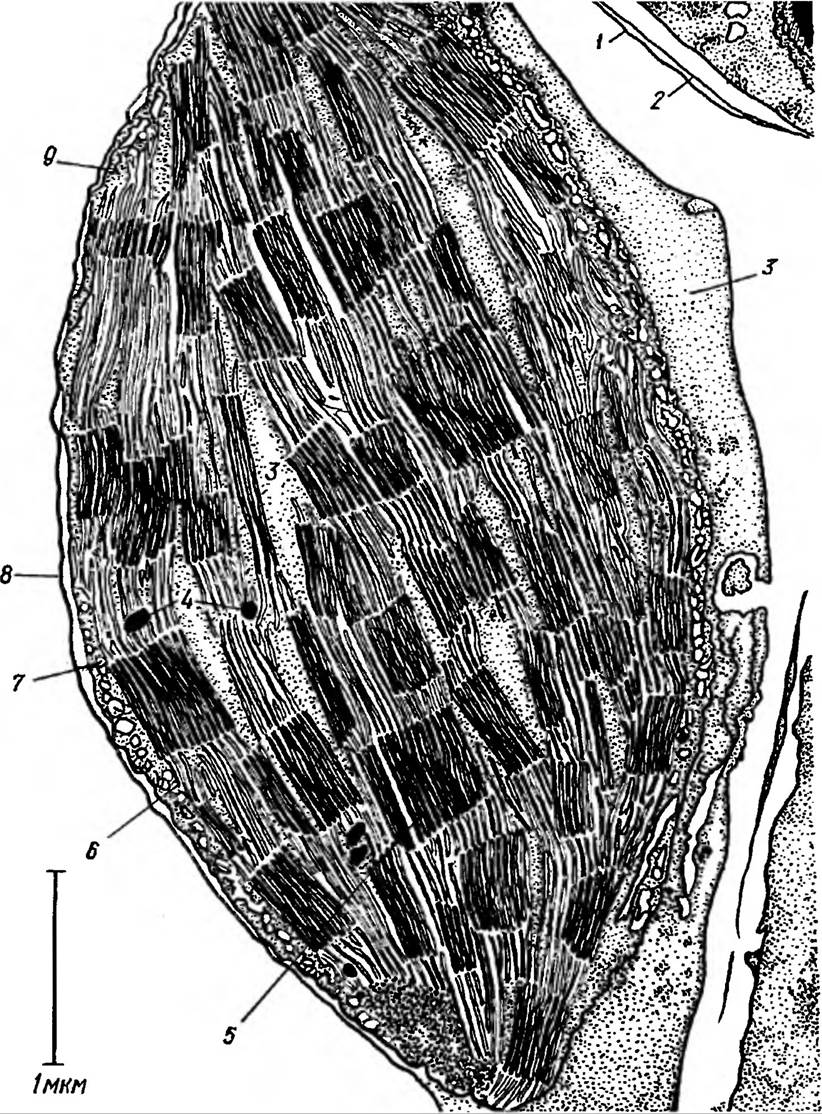
Рис. 37. Будова хлоропласта:
1 —зовнішня оболонка хлоропласта, 2 — внутрішня оболонка хлоропласта, 3 — строма, 4 — краплини ліпідів, 5 — грана, 6 — тилакоїди строми, 7 — крохмальні зерна, 8 — оболонка хлоропласта, 9 — Рибосоми
Кількість хлоропластів у клітині може коливатись від одного до 100 і більше. Загальна сумарна поверхня їх перевищує площу листків в десятки, навіть сотні разів.
Хлоропласти — це напівавтономна, саморегулююча система рослинної клітини, яка для свого розвитку та функціонування використовує генетичну інформацію як свого власного, так і ядерного, а можливо і мітохондріального геномів. У 1960 р. японський біохімік Івамура виявив в одноклітинній водорості хлорелі два типи ДНК — ядерну та хлоропластну. Стало очевидним, що генетичний матеріал евкаріот локалізований не лише в ядрі, яке служить головною базою збереження спадкової інформації, а й розсіяний по клітині в її субклітинних структурах.
Хлоропластна ДНК у вигляді подвійної спіралі має лінійну, кільце- та петлеподібну форми. Загальна кількість ДНК в хлоропласті залежно від виду коливається в межах 20-10-10-15 г, а молекулярна маса 84-104 мегадальтон (МД). Контурна довжина кільцеподібних молекул хлоропластної ДНК у вищих рослин досягає 43-55 мкм. Передбачають, що в такій ДНК знаходиться близько 2-Ю7 пар нуклеотидів, що відповідає наявності кількох тисяч генів.
Однак генетична інформація пластид досить обмежена. Тому багато компонентів пластид синтезуються під контролем як хлоропластної, так і ядерної ДНК. В хлоропластах знаходиться своя власна рибосомальна рРНК., а можливо, і повний набір своїх тРНК, які беруть участь в експресії геному організму. В стромі хлоропласта локазовані дрібні рибосоми з коефіцієнтом седиментації 70S. Маючи власний спадковий апарат, власну білоксинтезуючу систему, хлоропласт може контролювати синтез певних поліпептидів.
За хімічним складом основна маса хлоропластів складається з (% на суху речовину): білків (30-60), ліпідів (20-40), хлорофілів (5-9), каротиноїдів (4-5), ДНК (0,01-0,02), РНК (0,5-3,5), мінеральних речовин (6-10).
В них зосереджено до 80% всього заліза рослинної клітини, 65-70% всього цинку та близько 50% міді. Хлоропласти містять різноманітні Ферменти, які забезпечують функціонування ензиматичних реакцій фотосинтезу. В них знаходяться всі ферменти, які беруть участь в процесі фотосинтезу (рибулозобісфосфаткарбоксилаза — оксигеназа, окислювально-відновлювальні, синтетази, гідролази та інші).
Одним з найважливіших компонентів фотосинтетичного апарату є пігментний комплекс. Прокаріоти не мають спеціалізованих фотосинтетичних органел. Пігментний апарат їх представлений окремими мембранами, які дифузно розсіяні в клітині. Всі інші представники евкаріотів мають спеціалізовані Органели — хлоропласти, де і зосереджено весь пігментний комплекс. Як виявилось, фізико-хімічні властивості пігментів найкраще реалізуються саме в хлоропластах.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.