Фізіологія рослин - Мусієнко М.М. 2001
Хімічний та молекулярний склад, структура і функції рослинної клітини
Надходження іонів в рослинну клітину. Пасивне і активне надходження
Із довкілля в клітину рослин неперервно надходить не лише Вода, а й розчинені в ній солі. Надходження солей відбувається у вигляді іонів, які накопичуються у клітинній стінці, цитоплазмі, вакуолі.
Першим бар’єром на їх шляху виступає Клітинна стінка. Вона визначає не лише розмір і форму клітин рослини, а й її іонний баланс. Товщина клітинної стінки варіює від десятих долей мікрона до десятків мікронів. Мікрофібрили целюлози, які є її основними структурними одиницями, орієнтовані безладно. Вони переплітаються між собою в первинній стінці та розміщуються паралельно — у вторинній, створюючи проміжки до декількох десятків нанометрів в поперечнику. Ці проміжки, частково заповнюються водою, внаслідок чого із зовнішнього розчину можуть просуватися до плазмалеми, наявні у воді іони.
Подальший потік молекул та іонів в клітину контролюється плазмолемою. Транспортні процеси в плазматичній мембрані регулюють об’єм клітини, підтримують певний іонний склад гіалоплазми, що забезпечує сприятливі умови для прояву активності ферментів. Вони створюють іонні градієнти, необхідні для формування мембранного потенціалу тощо. Не дивлячись на обмежену проникність плазмолеми, Клітина постійно поглинає іони, про що свідчить розглянуте вище явище плазмолізу. В розчинах різних солей, як свідчать досліди М.Г. Холодного, форма плазмолізу різна. Кальцій підвищує в’язкість протоплазми, і, як наслідок, форма плазмолізу в розчинах його солей ввігнута. Калій знижує в’язкість — форма плазмолізу випукла. Разом з тим наявність плазмолізу показує, що у вакуоль солі якщо й проникли, то в незначній кількості.
Мембранне транспортування іонів може бути пасивним та активним. Пасивне поглинання(неспецифічна дифузія СО2 та О2, специфічні транслокатори, пори, канали, везикулярний транспорт) не вимагає витрати енергії. Воно пов’язане з процесом дифузії і відбувається за градієнтом даної речовини. З термодинамічної точки зору напрямок дифузії визначається хімічним потенціалом речовини. Для іонів напрямок дифузії визначається двома факторами: їх концентрація та електричний заряд. Чим вище концентрація речовини, тим вище її хімічний потенціал. Перенесення іонів проходить в бік меншого хімічного потенціалу.
Зауважимо, що напрямок руху іонів визначається не лише хімічним, а і електричним потенціалом. Наприклад, якщо іони надходять за градієнтом концентрації, то завдяки різній проникності мембрани швидше проникають або катіони, або аніони. Як результат, на мембрані виникає електричний потенціал, що, в свою чергу, викликає дифузію протилежно зарядженого іону. Таким чином, рушійною силою пасивного Транспортування іонів через мембрану є електрохімічний потенціал. Перенесення неелектролітів обумовлене градієнтом хімічного потенціалу, а перенесення іонів — градієнтом електрохімічного.
Пасивний транспорт може здійснюватися також завдяки полегшеній дифузії за участю специфічних переносників для певних іонів, але без додаткових витрат енергії. Завдяки переносникам, транспортування іонів проходить швидше, ніж при звичайній дифузії, звідси і назва — полегшена дифузія (рис. 17). Вважають, що полегшену дифузію обумовлюють білкові компоненти мембрани, які володіють здатністю «розпізнавати» певні речовини і переносити їх через мембрану.
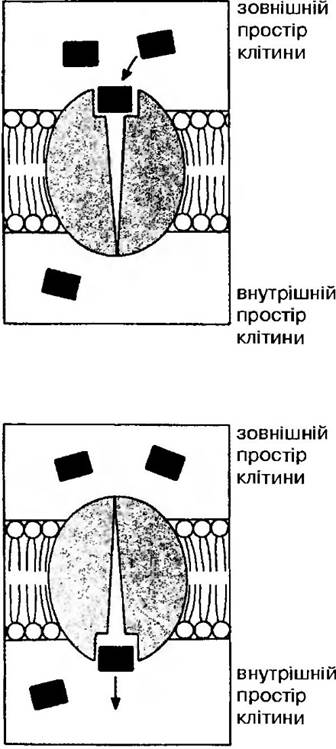
Рис. 17. Полегшена дифузія як спосіб перенесення речовин
Білки — транслокатори (переносники) часто розглядають як трансмембранні білки, які зазнають зворотніх конформаційних змін при перенесенні молекул через мембрану. Субстрат звязується із специфічним білком, переноситься через мембрану і звільняється. Всі транслокатори — інтегральні білки, які занурені в ліпідний шар мембран. Вони нерозчинні у воді, бо в їх складі переважають гідрофобні амінокислоти, такі як аланін, валін, лейцин, ізолейцин, фенілаланін. Завдяки різноманіттю та специфічності білків вони забезпечують вибірковість перенесення іонів.
Найбільш поширеними транспортними білками у рослин є транслокатори тріозофосфату та фосфору неорганічного у мембранах хлоропластів. Відомо, що цей транслокатор має дві субодиниці, кожна з яких складається із 324 амінокислотних залишків. Вони формують в мембрані хлоропласта 12 спіральних завитків, які перетинають товщу мембрани і забезпечують транспортний експорт фотоасимілятів із хлоропластів з швидкістю 80 молекул за секунду. Транслокатори функціонують у більшості мембран не лише рослинних клітин, а і у бактерій, тваринних організмів. Вони переносять різноманітні субстрати із швидкістю 10-10000 молекул за секунду.
Йонні канали — переносять різні іони через мебрану із швидкістю 106 -108 іонів за секунду. На відміну від транслокаторів, йонні канали відкриті з двох сторін мембрани. Рослини мають Н+, К+, Са2+, С- канали, а також для дикарбонових кислот (наприклад, малату) і, на відміну від тварин, неспецифічний канал для іонів Na+. Відкриття каналів регулюється електричним мембранним потенціалом, зв’язуванням його білків з Са++, або через їх фосфорилування. Виявлено, наприклад, що у продихових клітин, гіперполяризація плазматичної мембрани (100 мВ) відкриває канал для потоків іонів К+ всередину клітини, а деполяризація відкриває інші канали, якими К' покидає клітину. Тобто є два калієві канали — один спрямовує потік іонів всередину в Цитозоль клітини, інший — назовні. Основна структура йонних каналів — трансмембранні білкові спіралі, як і у транслокаторів.
Зовнішня мембрана хлоропластів, мітохондрій, пероксисом має в своєму складі білки — поріни, які представляють родину пори-формуючих білків (рис. 18). Поріни складаються із субодиниць масою 30 кД. Часто в мембрані вони знаходяться у вигляді тримерів, кожна субодиниця якого формує пору. Вони також формують трансмембранні спіралі із своїх амінокислотних залишків. Гідрофобні залишки амінокислот спрямовані в ліпідний шар, а гідрофільні — в білковий шар мембрани. В порівнянні з білками іонних каналів поріни мають більш економну структуру, так як формуються однією молекулою поріну. Вони формують пори для дифузії деяких субстратів і води.
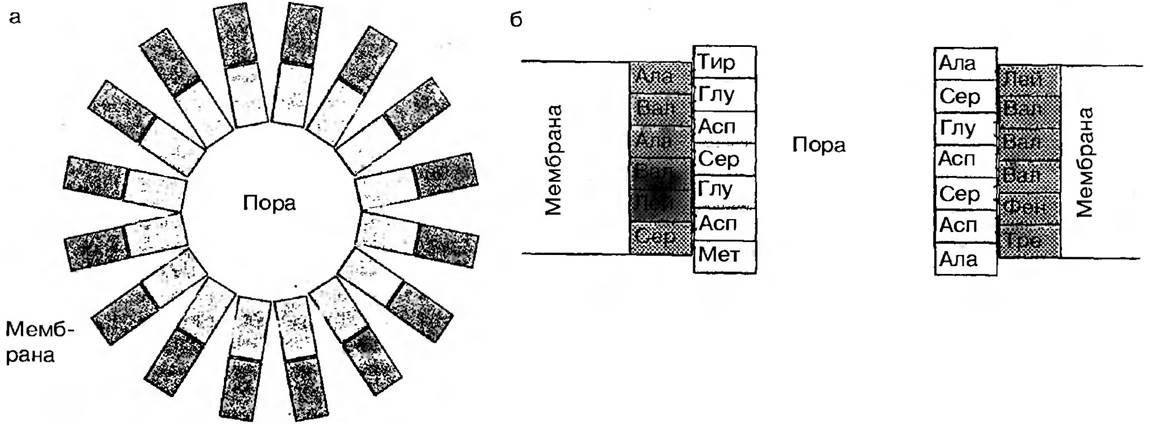
Рис. 18. Діаграма структури пори мембрани, сформованої порінами
а — вид зверху, б — поперечний розріз
Водні канали забезпечують перенесення води через мембрану. Як виявилось недавно, канали формує родина однакових для різних організмів білків — агвапорінів (aguaporins). Канали складаються із 6 трансмембранних спіралей, які формують в мембрані тетрамер, де кожний мономер слугує окремим каналом, через який щосекунди транспортується 10 9-1011 молекул Н2О.
Активний транспорт — це поєднаний з використанням енергії процес перенесення молекул або іонів через мембрану проти електрохімічного градієнта. Енергія необхідна тому, що речовина повинна рухатися всупереч своєму природному прагненню дифундувати в протилежному напрямку. Він вибірково концентрує іони в клітині і має вирішальне значення для її життєдіяльності. Енергія, в основному, акумульована в АТФ і звільняється при гідролізі її за участю фермента аденозинтрифосфатази (АТФ-аза).
Для рослинної клітини велике значення має Н+-АТФ-аза (водневий насос, або протонна помпа), яка забезпечує перенесення протонів через мембрани, використовуючи енергію гідролізу АТФ. Цей процес супроводжується спряженим оберненим перенесенням катіонів через мембрану (антипорт). Одночасно з іоном водню Н+ в той же бік можуть переноситись аніони (симпорт) (рис. 19). Допускають, що АТФ-аза розташована впоперек мембрани. Звільнена при
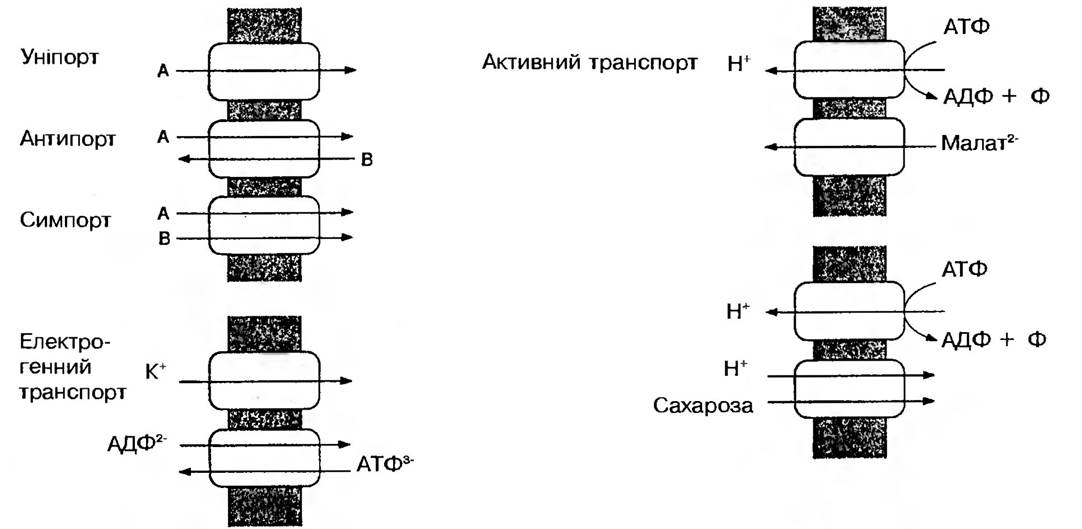
Рис. 19. Класифікація мембранних транспортних процесів
гідролізі енергія АТФ змінює конфігурацію самої АТФ-ази, завдяки чому ділянка фермента, яка звязує певний іон, повертається і виявляється по інший бік мембрани. Можливо також, що переносниками виступають специфічні білки, а енергія АТФ, що звільняється за участю АТФ-ази, використовується на їх переміщення через мембрану. Розрахунки показують, що для того, щоб 1 моль солі дифундував проти градієнта концентрації, необхідно витратити близько 4600 Дж. Як відомо, під час гідролізу АТФ виділяється 30660 Дж/моль, чого досить для транспорту декількох молей солі.
Нарешті, клітина здатна "поглинати" певні речовини разом з водою шляхом піноцитозу. Спочатку проходить адсорбція іонів на певній ділянці плазмолеми, далі її упинання, утворення пухирців із рідиною, які пізніше зливаються з мембранами ендоплазматичного ретикулуму або іншими мембранами.
Після того як молекули пройдуть бар’єр клітинної стінки та плазматичної мембрани, вони можуть відносно швидко розподілятися по всій клітині шляхом дифузії, а також завдяки руху цитозоля. Істотна роль в процесі зв’язування певних іонів цитоплазмою належить клітинним ор- ганелам, зокрема, хлоропластам і мітохондріям.
У вакуоль попадають іони у випадках, коли вони вже не потрібні для протоплазми. Щоб попасти у вакуоль, іони повинні подолати ще одну перепону — тонопласт. Перенесення іонів через тонопласт також відбувається з допомогою переносників і в більшості випадків вимагає витрат енергії. Прониклі у вакуоль речовини забезпечують осмотичні властивості клітини.
Таким чином, прониклі через клітинну стінку і плазмалему іони накопичуються і включаються в метаболізм в цитоплазмі, і лише їх надлишок дифундує до вакуолі. Саме тому не буває рівноваги між вмістом іонів у зовнішньому розчині та їх наявністю у клітинному соку. Завжди, чим швидше включаються іони в обмін речовин, тим інтенсивніше проходить їх поглинання.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.