Фізіологія рослин - Мусієнко М.М. 2001
Загальні принципи регуляції росту та морфогенезу рослин
Генетична регуляція
Генетична регуляція забезпечує:
·збереження та реплікацію Інформації про Індивідуальну будову конкретного організму І послідовність процесів морфогенезу, які зашифровані в хроматині ядра та кільцевих ДНК пластид, мітохондрій у вигляді триплетного коду;
·регулює процес зчитування (транскрипцію) даної Інформації, процесинг синтезованих РНК І їх транспорт, збереження мРНК в латентній формі у вигляді Інформосом та їх активацію;
·контроль за збиранням рибосом, полірибосом та відповідним синтезом поліпептидів (трансляцію).
РІСТ І РОЗВИТОК регулюються насамперед на клітинному рівні. Ріст органа або організму відповідно складається з росту його клітин, а морфогенез, тобто утворення специфічних форм організації органа чи організму, є результатом тих шляхів розвитку, на які стають окремі клітини. Детермінація шляхів розвитку кожної клітини є основою фізіології розвитку. При детермінації Клітина робить вибір із численного набору потенціальних можливостей (генів, інформацій), хоча більша частина інформації все ж залишається незадіяною (зокрема, гени, що забезпечують виконання основних функцій клітини — синтез білка, ГЛІКОЛІЗ тощо).
Ембріональна меристематична клітина може розвиватися різноманітними шляхами. При цьому можуть зникати певні Органели клітини, наприклад клітинне Ядро та тонопласт у члеників ситоподібних трубок, або навіть зникає вся протоплазма (членики судин). Може збільшуватися кількість мітохондрій (клітини-супутники), можуть розчинятися клітинні стінки (ксилема), в клітинних стінках може відкладатися лігнін або суберин. Нарешті, з пропластид можуть утворюватись хлоро-, лейко- і хромопласти.
Перехід на той чи інший шлях розвитку визначається особливим набором білків, тобто кожна спеціалізована клітина синтезує лише частину генетично можливих ферментів. Так, висока концентрація амілази спостерігається лише в алейроновому шарі зернівок, фенолоксида- зи містяться в ендодермі, а не в корі кореня і т. д.
У зв’язку з тим що синтез кожного білка контролюється через матричну ДНК одним Геном, утворення різних наборів ферментів в різних типах клітин, можливо, е результатом диференціальної активації генів (активація однієї групи генів при одночасній інактивації інших).
У процесі формування, наприклад сім’ядолі, при постійній структурі ДНК послідовно змінюється склад РНК (склад азотистих основ), а значить і білків (електрофоретичні властивості, ферментативна активність). Така біохімічна диференціація є не результатом, а причиною морфологічного диференціювання. Диференціальна активація генів відбувається не лише в спеціалізованих ембріональних клітинах. Наприклад, світло може спричинити утворення волосків або продихів в клітинах епідерми — це процеси, для яких також потрібна диференціальна активація генів.
Морфогенетична РНК. Матричну РНК, яка утворюється завдяки диференціальній активації генів і яка бере участь у морфогенез!, називають морфогенетичною РНК Роль даної РНК легко простежити на прикладі водорості ацетабулярії (рис. 170).
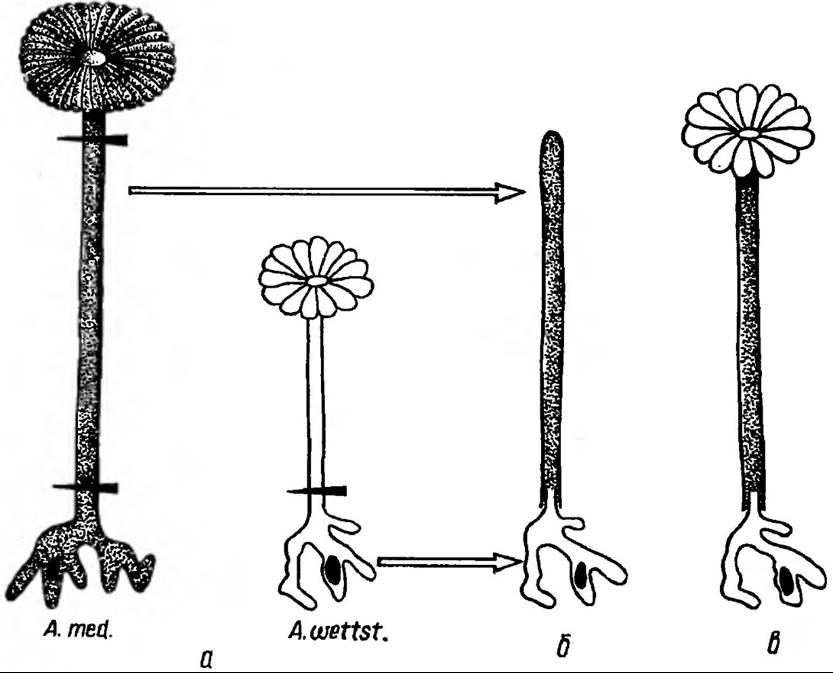
Рис. 170. Доказ наявності морфогенетичних речовин у ацетабулярії: а — відрізок ніжки (без’ядерний) А. mediterranea зщеплений на ризоїд (містить ядро) A. wettsteinii; б — компоненти зщеплення зростаються; в — відрізок ніжки A. mediterranea регенерує шапинку A. wettsteinii
Рослини її досягають до 5 см, розміри ядра до 0,1 мм. Це одноклітинна водорість, яка має ризоїд, ніжку і у різних видів різну за формою шапинку. Якщо за допомогою трансплантації ядра генетичний матеріал перенести із ацетабулярії одного виду в молоді, які ще не мають шапинки, ацетабулярії іншого виду, у яких попередньо видаляють власне ядро, формується вид шапинки, характерний для рослини-донора ядра, тобто ядро визначає форму шапинки, яка утворюється. Значить, у клітинному ядрі утворюється специфічна для кожного виду морфогенетична матрична РНК, яка накопичується апікально, в сторону верхівки, і там керує морфогенезом утворення шапинки через синтез специфічних білків. Тому немає сумнівів, що і в клітинах вищих рослин така морфогенетична РНК може виступати посередником між вибірковою активацією генів та клітинною диференціацією.
Включення диференціальної активації генів. До факторів, які зумовлюють вибіркову активацію генів, належать фітогормони та світло. Досить часто для збудження однієї реакції треба кілька сигналів, наприклад дії двох фітогормонів. На той самий сигнал не всі клітини реагують однаково. Наприклад, у відповідь на сигнал довгий темновий період Тканини конуса наростання пагона будуть переходити до формування квіток, чого не спостерігається у конуса наростання кореня. Ще один приклад, коли під впливом світла лише клітини епідермісу здатні утворювати волоски або продиховий апарат. Це свідчить про те, що вирішальним є не стільки діючий фактор, скільки генна модель тієї клітини, що його приймає, а саме набір регульованих генів, що є в розпорядженні даних клітин. Згадайте, що геном клітини в кожний момент містить активні, блоковані та регульовані гени. Тільки регульовані гени включаються або виключаються при диференціальній активізації, причому одні фактором А, інші — В, С і т.д. Можливі також випадки, коли відбувається не активація генів, а безпосередньо активація уже сформованих білків-ферментів.
Координована активація генів. При детермінації розвитку емріональної клітини беруть участь різноманітні гени. Тому важко собі уявити, що певний фактор, наприклад гормон, окремо активує кожний ген, або кожну групу генів, тому що при впорядкованому розвитку гени повинні ставати активними один за одним. Тому можна припустити, що гени об’єднані в функціональні одиниці, які можна порівняти з програмами, закладеними в ЕОМ. Геном містить обмежену кількість програм, які можуть бути запущені відповідними факторами. Як і в ЕОМ, до складу програми входять підпрограми, підпідпрограми і т. д.
Розглянемо приклад програми: палісадні клітини мезофілу → формування хлоропластів → синтез хлорофілів і т. ін. Інша програма: квітка → підпрограми → тичинки, пилок і т. д. Деякі підпрограми належать лише до однієї програми (тичинка —до квітки), інші — до кількох (ксилема належить до пагона і до кореня). Відповідно для кожної такої генетичної програми існує конкретна молекулярна конструкція.
Якщо геном складається з невеликої кількості таких елементів, то вони можуть бути взаємозв’язані завдяки тому, що вони крім звичайних генних продуктів (мРНК) виробляють ще й молекули-модулятори, функцією яких є включення або виключення інших елементів геному. Молекули модуляторів, можливо, також складаються з РНК або є вторинними генними продуктами — білками. Існування такого роду взаємозв’язків пояснюють явища індукції та репресії.
Є також дані про наявність елементів геному, що утворюються як єдиний продукт молеку- ли-модулятора і є виключно для координованої активації генів. Такі елементи, які складаються з інтегрованих генів, називають інтеграторами. Принцип їхньої дії можна уявити так: фактори, які включають ген, можуть діяти, можливо, через проміжний блок на молекули модуляторів або на особливі сенсорні гени, які контролюють дію інтеграторів так само, як ген — оператор — Оперон. Ці моделі дають змогу пояснити два відомі факти: по-перше, у вищих рослин є численні повтори послідовностей ДНК — це і є, можливо, гени-інтегратори та рецептори. По-друге, більша частина м-РНК руйнується в ядрі — тому це, можливо, РНК, яка утворена генами- інтеграторами.
Слід пам’ятати, що в процесі розвитку гени активуються один за одним у певній послідовності, причому детермінація обмежується, як правило, першими кроками процесу розвитку. Подальші зміни генної моделі координуються внутрішніми механізмами.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.