Фізіологія рослин - Мусієнко М.М. 2001
Кореневе живлення рослин
Фізіологічна роль металів-макроелементів
Калій
Більшість рослин характеризується високим вмістом калію (0,9-1,2%), хоча дуже незначна частина його знаходиться в клітинних структурах у зв’язаному стані. Це типово мобільний елемент. Калій рослини одержують із розчинених в ґрунтовому розчині його солей (0,5-2% валового запасу в грунті). Найбільшим вмістом характеризуються меристеми, молоді пагони, листки, бруньки. В рослинній клітині близько 80% його міститься в вакуолях. Більша частина його (70%) в клітині перебуває у вільній іонній формі, решта (30%) в адсорбованому стані. Калій не входить до складу жодної органічної сполуки. Поглинається у вигляді катіону К+. Калій — осмотично активний елемент, сприяє гідратації протоплазми, знижує її в’язкість та підвищує оводненість.
Роль калію в фотосинтезі пов’язана з його участю в регуляції роботи продихового апарата, а також фотофосфорилування. Вважають, що тургор клітин змінюється як наслідок функціонування калієвих іонних насосів. Відкриті продихи характеризуються високим вмістом калію в замикаючих клітинах, при закриванні їх іони калію викачуються назовні.
Зменшення вмісту калію до 0,2-0,6% зумовлює різке зниження інтенсивності фотосинтезу, гальмує ріст, порушує фосфорний метаболізм, синтез пігментів, білків, вуглеводів.
Перерозподіл фотоасимілятів пов’язаний також із вмістом калію в провідних клітинах флоеми, де він переважно локалізується. Вважають, що загальний осмотичний рівень в ситоподібних трубках підтримується шляхом взаємної компенсації сахарози та іонів калію. Катіонна рівновага в них зсунута в бік іонів калію, що є передумовою для розвитку високих електрохімічних градієнтів на мембранах та для активного функціонування N+-К+-насосів.
Оптимальний вміст калію забезпечує баланс процесів синтезу та гідролізу в клітині. Саме іони калію створюють іонну асиметрію, а отже, мембранний потенціал між клітиною та середовищем.
Калій — активатор понад 60 ферментних систем, але не входить до них як структурна частина. Він активує включення фосфатів в Органічні сполуки, бере участь в синтезі рибофлавіну — компоненту всіх флавінових дегідрогеназ. Стимулюючи синтез крохмалю, сахарози, моноцукрів, пектинових речовин, він найбільше сприяє формуванню високих якісних показників всіх плодових. Між іншим, для формування якості плодів надлишок калію інколи шкідливіший, ніж його дефіцит, бо в таких умовах послаблюється ефективність кальцію та магнію.
Властивість іонів калію підтримувати сприятливі для життя цитоплазми відповідні фізико- хімічні особливості колоїдів — еластичність, в'язкість, дисперсність, оводненість має надзвичайне значення в формуванні стійкості рослин до несприятливих умов середовища.
Для нього характерна реутилізація, під кінець вегетації можна спостерігати міграцію калію із старіших частин рослинного організму до молодших. Його дефіцит спричинює зниження домінуючого ефекту апікальних бруньок.
Критичний період в потребі даного елемента припадає на перші стадії розвитку. Дефіцит його спричинює пожовтіння листків нижнього ярусу, побуріння країв решти листя, ніби від запалення. У зв’язку з тим, що цей елемент специфічно впливає на цитоплазму, його не можна замінити іншим, хоча деякі фізіологічні процеси замінюються натрієм та рубідієм. Калійні добрива особливо сприятливо впливають на врожайність пшениці, цукрових буряків, картоплі та плодових культур.
Кальцій
Вміст кальцію в рослинах до 0,2%, тоді як в старих листках його кількість досягає 1% Надходить в рослинний Організм у вигляді іонів Са2+. Щодо кальцію рослини поділяють на кальціє-
філи (боби, нут), кальцієфоби (люпин жовтий, кукурудза), які практично не мають вільного кальцію, та нейтральні види (гарбуз).
Більшість хрестоцвітих, бобових поглинають іони кальцію в значних кількостях та нагромаджують його в клітинному соку. Менше його в зернових культурах, цукровому буряку. З віком все більше його накопичується в листках та інших вегетативних органах. У процесі Старіння клітини кальцій із цитоплазми переходить у вакуолі, де відкладається у вигляді різноманітних кристалічних включень солей органічних кислот. В насінні він зустрічається в складі фітину.
Роль кальцію різноманітна. Кількісно основна функція кальцію полягає у включенні його в структуру серединної пластинки клітинної стінки, де він виступає як обмінний фонд кальцію. Зв’язуючись з пектинами, він утворює нерозчинні солі пектатів кальцію, що спричинює затвердіння напіврідкої структури клітинної стінки. Серединні пластинки, що склеюють клітинні стінки сусідніх клітин, складаються переважно із пектатів кальцію. При дефіциті кальцію формування нових клітинних стінок припиняється, хоча поділ ядра відбувається нормально.
Кальцій, на противагу калію та магнію, міцно зв’язується з різними органічними сполуками в клітині, тому він малорухомий, хоча є дані, що він здатний до повторного використання рослиною.
Кальцій — компонент багатьох органел клітини (хлоропласти, Мітохондрії, Рибосоми, Ядро). Він підтримує колоїдно-хімічні властивості протоплазми, впливає на гідратацію її, в’язкість. Він може регулювати рух цитоплазми, впливаючи на структурні компоненти цитоскелету — мікротрубочки. Кальцій також потрібний для гранулокриновоі' секреції компонентів клітинних стінок за участю диктіосом апарата Гольджі. Завдяки впливу на процес формування елементів цитоскелета, він також пов'язаний з мітозом. Кальцій реагує з різними органічними кислотами, утворюючи солі, завдяки чому виступає регулятором pH клітинного соку.
На сьогодні можна стверджувати, що іони Са2+ впливають на різні види внутрішньоклітинної активності. Він активатор деяких ферментних систем (фосфорилази, α-амілази, глутамат- гідрогенази, глюкозо-6-фосфатдегідрогенази, ліпази, аденозинтрифосфатази).
Іони кальцію виявлені в зоні ядра, що, можливо, пов’язане з потребою його для процесів клітинного поділу та росту в фазу розтягування. Проростання пилку та ріст пилкової трубочки стимулюється іонами даного елемента. Підвищені дози кальцію гальмують ауксинезалежний ріст при одночасному підсиленні здатності ауксинів до індукції електрофізіологічної, поляризації клітин.
Як відомо, пектини та інші сполуки клітинної оболонки містять значну кількість карбоксильних груп, які при ДИСОЦІАЦІЇ відщеплюють водень, що надає в цілому клітинній оболонці від’ємний заряд. Вони електростатично притягують іони кальцію (Са2+), тобто постійно відбувається обмін Н+ на Са2+. Майже вся катіоннообмінна ємність кореневої системи зайнята Са2+ та Н+, тому даний елемент відіграє важливу роль в процесах первинного поглинання елементів мінерального живлення.
Слід зауважити, що іони Са++ впливають на різні види внутрішньоклітинної активності через їхній зв’язок із специфічним білком кальмодуліном. Він широко відомий у тваринному та рослинному світі, його молекулярна маса 16000-19000. Майже в усіх живих істот кальмодулін має ту саму амінокислотну послідовність, насичений залишками глутамінової та аспарагінової кислот. В еволюційному плані це один з найдавніших і дуже консервативний білок. Виступаючи в ролі кальцієвого внутрішньоклітинного рецептора, він регулює в клітині більшість Са2+- залежних процесів (активність протеїнкіназ, транспортних Са2+-АТФаз, концентрацію внутрішньоклітинного кальцію, участь в секреторних процесах) (рис. 137). Кальмодулін в стані насичення його іонами Са2+ надзвичайно термостабільний. Є дані про зв’язок його з гравітро- пічною реакцією кореневого чохлика.
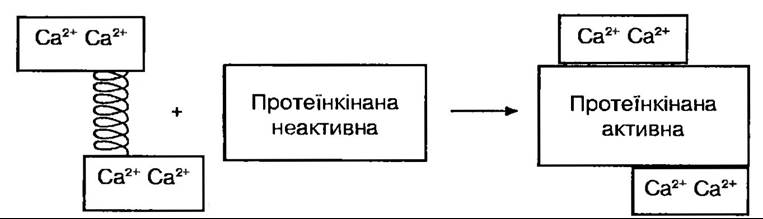
Рис. 137. Активування протеїнкіназ Са2+ кальмодуліном
Дефіцит кальцію впливає насамперед на меристеми і кореневі системи. Під час поділу клітин пригнічується утворення клітинних стінок, припиняється утворення бічних коренів, кореневих волосків. У молодих листків по краях з’являються світло-зелені смуги, Тканини деяких ділянок відмирають (некроз), верхівкові бруньки часто теж відмирають, а кінчики листочків закручуються вгору. Некротичні плями з’являються також на плодах і в запасаючих тканинах. Характерні зміни процесу обміну, пов’язані з дефіцитом кальцію, виявляються в зміні напрямку синтезу вуглеводів та азотистих речовин.
Магній
Вміст даного елемента в рослинах залежно від виду, фізіологічного стану та умов живлення коливається в межах від 0,013 до 3,15% на суху речовину. Надходить в рослину у вигляді іонів Мд2+. Магній входить в склад основного пігменту зелених листків — хлорофілу. Він накопичується переважно в найбільш життєздатних тканинах — меристемах, вузлах кущіння, зародку зернівок. З початком плодоутворення він нагромаджується в насінні, утворюючи там разом з фосфорною кислотою фітин. Він досить рухомий, що зумовлює його деяку реутилізацію із старіючих органів. Найбільший процентний вміст магнію в пластидах, мітохондріях, пектатах первинної клітинної стінки.
Дія магнію на рослини характеризується високою різноманітністю. Він може впливати на зміну просторової структури білкових молекул в бік відщеплення асиметрії їх. Це одна з причин його впливу на зміну структурної в’язкості протоплазми та на рухомість внутрішньоклітинної води.
Та особлива функція магнію пов’язана з участю його в фотосинтезі як компонента хлорофілу, який містить до 15% всього вмісту в рослині. Він є не лише складовою частиною хлорофілів, а й бере участь в початкових стадіях біосинтезу порфіринового ядра, зокрема в синтезі протопорфіну IX. Магній регулює структуру органел, збільшуючи активність первинних процесів фотосинтезу. Вважають, що він впливає на формування з окремих фотохімічних реакційних центрів та світлозбираючих комплексів, крупніших мультицентральних фотосинтетичних одиниць. Він активує транспортування електронів між двома фотосистемами, фотофосфорилування, відновлення НАДФ+.Збільшення концентрації магнію в стромі хлоропласта активує рибулозобісфосфаткарбоксилазу, підвищує спорідненість її до СО2.
Іони магнію відіграють важливу роль в функціонуванні білок-синтезуючої системи. Вони підтримують цілісність структури рибосом, звязуючи РНК і білок. Велика і мала субодиниця рибосом асоціюються лише в присутності іонів магнію. Дефіцит іонів магнію не лише спричинює дисоціацію часточок SOS-рибосом на 30S та 50S, а й розкручування ще менших фрагментів. По-друге, магній сприяє закріпленню на рибосомах, матричної та транспортної РНК за рахунок електростатичного зв’язку з Мg2+. Іони даного елемента поліпшують умови включення амінокислот в білки. Він активує ДНК та РНК-полімерази, бере участь в формуванні певної просторової структури нуклеїнових кислот.
Функціонування багатьох ферментів пов’язане з участю магнію, причому найбільш значною самостійною групою є фосфотрансферази, для яких Mg-АТФ може виступати як субстрат (фос- фокінази, АТФ-ази, пірофосфатази). До ферментів, які специфічно активуються магнієм, належать 5-нуклеотидаза, аргінінсукцинатсинтетаза, рибулозобісфосфаткарбоксилаза, альдолаза та ін. Більшість реакцій гліколізу, циклу Кребса активується магнієм. В ряді випадків вплив магнію на роботу ферментів визначається тим, що він реагує з продуктами реакції, зміщуючи рівновагу в сторону їх утворення.
Зв’язок магнію з активністю ферментів визначає роль його в метаболізмі рослин, в процесах росту й розвитку.
Активація магнієм нітратвідновлюючої здатності рослин та одного з ключових ферментів асиміляції азоту — глутамінсинтетази відіграє позитивну роль в азотному обміні рослин. Він спричинює значні зміни і в фосфорному обміні, що обумовлене роллю магнію в активації ферментів фосфорного метаболізму.
Магній здатний підсилювати синтетичні процеси, поєднані з утворенням різних ефірних олій, вітамінів, каучуку.
Зовнішні ознаки дефіциту магнію — жилки листків залишаються зеленими, а жовкне лише пластинка між ними. Магнієвий хлороз дістав назву мармурогюдібного хлорозу листків. Дефіцит настає, якщо вміст магнію в грунті менший 2 мг на 100 г грунту. Особливе значення має магній для рослин, які розвиваються в умовах кислих грунтів.
З метою забезпечення рослин магнієм використовують калійно-магнієві та комплексні добрива, куди, входить калій, магній та фосфор.
Залізо
У грунті вміст заліза коливається від 1 до 7%, в рослині — 0,02 — 0,08%, тому воно знаходиться на межі між макро- та мікроелементами. Надходить в рослину у вигляді іонів Fe3+. Його роль пов’язана, насамперед, з його здатністю-легко переходити із окисленої-форми (Fe3+) у відновлену (Fe2+) і навпаки. Відсутність заліза або навіть нестача його спричиняє дуже помітні порушення функціональної діяльності рослин.
Залізо як елемент мінерального живлення бере участь в структурі окремих ферментів і цілих ферментних систем, пов’язаних з окислювально-відновними реакціями клітини. Крім процесів фотосинтезу, Дихання, його іони беруть участь у відновленні нітратів, біологічній азотфіксації, як складові компоненти нітратредуктази та нітрогенази.
Роль фередоксинів та цитохромів у обміні рослинної клітини визначається насамперед каталітичними властивостями заліза, що є конституційною та функціональною складовою частиною цих сполук. Досить згадати про значення в окислювальному та енергетичному обміні тканин, щоб стала зрозумілою та величезна роль, яку залізо виконував процесах життєдіяльності.
Прикладом участі заліза в біосинтетичних реакціях можуть бути реакції синтезу флавопротеїнових ферментів. Залізо необхідно для утворення хлорофілів. Воно каталізує утворення його попередників (амінолевулінової кислоти та протопорфиринів).
Залізо може знаходитись і в запасній формі у вигляді білка феритину, який містить залізо в негеміновій формі.
Дефіцит заліза найчастіше спостерігається на карбонатних грунтах, що можна пояснити переходом його в малодоступний для рослин стан. В таких умовах досить ефективне застосування комплексних органічних сполук заліза — хелатів. У тілі рослин залізо міститься у вигляді комплексних органо-мінеральних сполук, тоді як його Неорганічні сполуки відсутні. Залізо не підлягає реутилізації. Цим пояснюється той факт, що хлороз здебільшого властивий листкам верхніх ярусів. Якщо рослини не вбирають залізо протягом тривалого часу, то листки буріють, а потім відмирають. При недостачі заліза можна застосувати позакореневе живлення.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.