Фізіологія рослин - конспект лекцій - О. М. Тарнопільська 2019
4. Фотосинтез
4.5 Нециклічне й циклічне фотосинтетичне фосфорилювання
Фотосинтетичне фосфорилювання, тобто утворення АТФ у хлоропластах у процесі реакцій, що активуються світлом, може здійснюватися циклічним і нециклічним шляхами.
Циклічне фотофосфорилювання є простішим й еволюційно більш древнім. У разі циклічного фотофосфорилювання функціонує тільки ФСІ та її єдиним продуктом є АТФ.
Циклічне фотофосфорилювання було відкрито в 1954 р. Даніелем Арноном, Дж. M. Алленом і Альбертом Френкелем на ізольованих хлоропластах шпинату.
У фотосистемі II реакційний центр, що складається з димеру хлорофілу а з максимумом поглинання при 680 нм (П680), послідовно поглинувши енергію, що дорівнює двом квантам короткохвильового червоного світла, і перейшовши в синглетний збуджений стан, передає два електрони феофітину. Від феофітину електрони послідовно передаються на пластохинони QA і QB, розташовані на іншому боці мембрани тилакоїду, потім на дві ліпідорозчинні молекули пластохінона PQ. Він є переносником протонів і електронів, що виконує в мембрані роль човника. Беручи два електрони й захоплюючи зі строми 2 Н +, дві молекули PQH2 дифундують через ліпідну фазу мембрани. Електрони від 2 PQH2 надходять у ланцюг:
- залізосірчаний білок FeSr → цитохром f → білок, що містить Cu, пластоціанін → П700, а протони потрапляють у порожнину тилакоїда.
Вакантні місця в П+680 заповнюються двома електронами з переносника електронів, що містить Mn, який відновлюється за участю білкового комплексу S. Окислений комплекс S зв’язує воду та відновлюється за рахунок електронів води. Під час фотоокислення води комплексом S друга пара протонів звільняється в порожнину тилакоїда, а атом кисню дифундує з хлоропласта.
Після збудження реакційного центру фотосистеми I - молекули хлорофілу а з максимумом поглинання при 700 нм (П700) двома квантами довгохвильового червоного світла два електрони передаються мономерній формі хлорофілу а - А1 і потім переносникам електронів залізосірчаним білкам А2 і АВ, фероксідіну на зовнішньому боці мембрани тилакоїду, НАДФ-редуктази. Цей фермент відновлює НАДФ у НАДФН. На вакантні місця в П+700 переходять електрони з пластоціаніну, і нециклічний ланцюг транспорту електронів замикається.
У мембранах хлоропластів може функціонувати циклічний транспорт електронів, коли функціонує тільки фотосистема I. Збуджені молекули П700 передають електрони на А1, залізосірчані білки А2 і АВ, які переносять електрони на іншу сторону мембрани, ферредоксин, пластохінон, цитохром b6, залізосірчані білок FeSr, цитохром f, пластоціанін і П700 (рис. 4.4). Енергія, що звільняється під час транспортування електронів, використовується для фосфорилювання АДФ.
Синтез АТФ. Передача електрону по ЕТЛ проходить за градієнтом окисно-відновного потенціалу й супроводжується вивільненням енергії, яка використовується для синтеза АТФ. Синтез АТФ відбувається за допомогою електрохімічного градієнта протонів, що накопичуються у полості тилакоїду грани.
Існує декілька теорій, що пояснюють механізм фосфорилювання АДФ, поєднаний із транспортом електронів. Найвизнаннішою є хеміосмотична теорія англійського біохіміка П. Мітчела (1961). За цією теорією, пластохінон, приєднавши два електрони від П680, приєднує ще два протони зі строми хлоропласта та переносить їх крізь мембрану до внутрішньотилакоїдного простору. Протони накопичуються всередині тилакоїда і внаслідок цього відбувається фотоокислення води.
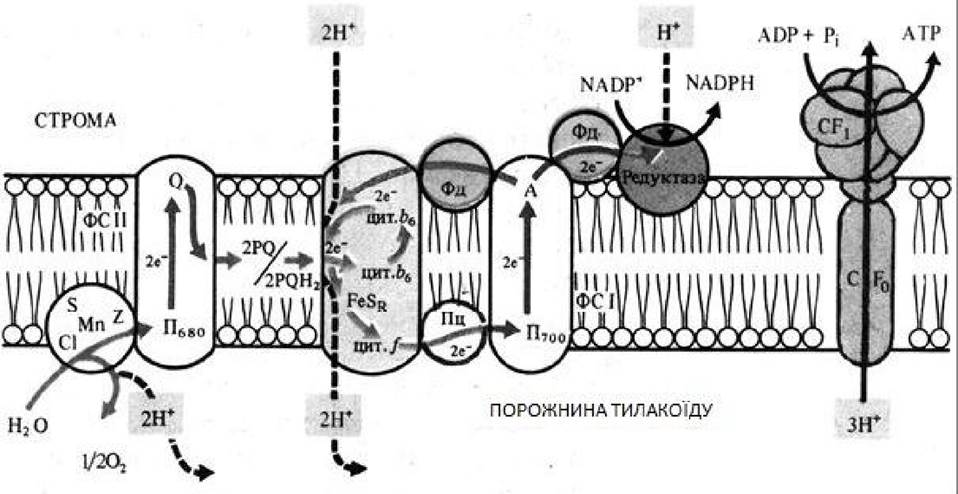
Рисунок 4.4 - Локалізація електрон- і протонтранспортних реакцій у тилакоїдній мембрані хлоропласта (за В. В. Польовим)
Завдяки нерівномірному розподілу протонів по обидва боки мембрани утворюється різниця хімічних потенціалів протонів, і виникає електрохімічний мембранний потенціал іонів водню (ΔμН+). ΔμН' включає два складники: концентраційний (ΔμН), що виникає внаслідок нерівномірного розподілу протонів по обидва боки мембрани, і електричний (Δψ), що обумовлений виникненням протилежного заряду на поверхні мембран, тобто утворенням мембранного потенціалу. Енергія ΔрН і Δψ використовується для зворотного транспорту протонів із внутритрішньотилакоїдного простору в строму хлоропласта по особливим каналам. Зі зворотним транспортом протонів спряжено фосфорилювання АДФ.
Спряження зворотного транспорту протонів і фосфорилювання АДФ забезпечує Н+-АТФ-синтетаза, що розташована в тилакоїдних мембранах і складається з двох частин: водорозчинної каталітичної частини (F1), зверненої до строми хлоропласта, і мембранної частини (F0), що пронизує подвійний шар ліпідів. Остання становить протонний канал, по якому протони можуть повертатись у строму хлоропласта. АДФ і фосфат з’єднуються з ферментом у його каталітичній частині F1. Два протона переміщуються по градієнту електрохімічного потенціалу по протонному каналу (F0), з’єднуються з киснем фосфату, утворюючи воду. Втрата кисню активує фосфатну групу, і вона приєднується до АДФ з утворенням АТФ.
Фермент Н+-АТФ-синтетаза активний, поки транспортуються протони. Протони пересуваються, якщо їхня концентрація у внутрішньотилакоїдному просторі більше. На кожні два електрони, переданих по електронтранспортному ланцюгу, усередині тилакоїду накопичується 4Н+. У разі повернення назад у строму хлоропласта двох протонів синтезується одна молекула АТФ.
Последнее обновление: 28/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.