Фізіологія рослин - Мусієнко М.М. 2001
Фотосинтез: фізіолого-біохімічні та екологічні аспекти
Фотоіндуковані окислювально-відновлювальні перетворення компонентів ЕТЛ та фотосинтетичне утворення АТФ і НАДФ×Н
Фотохімічні реакції фотосинтезу — це реакції, в яких енергія світла перетворюється в енергію хімічних звязків, і в першу чергу в енергію фосфорних звязків АТФ. Нагадаємо, що саме АТФ є енергетичною валютою клітини, яка забезпечує протікання всіх процесів в ній. Молекула хлорофілу реакційного центру, віддаючи електрон акцептору, окислюється. Електрон надходить в електрон- транспортний ланцюг (рис. 63).
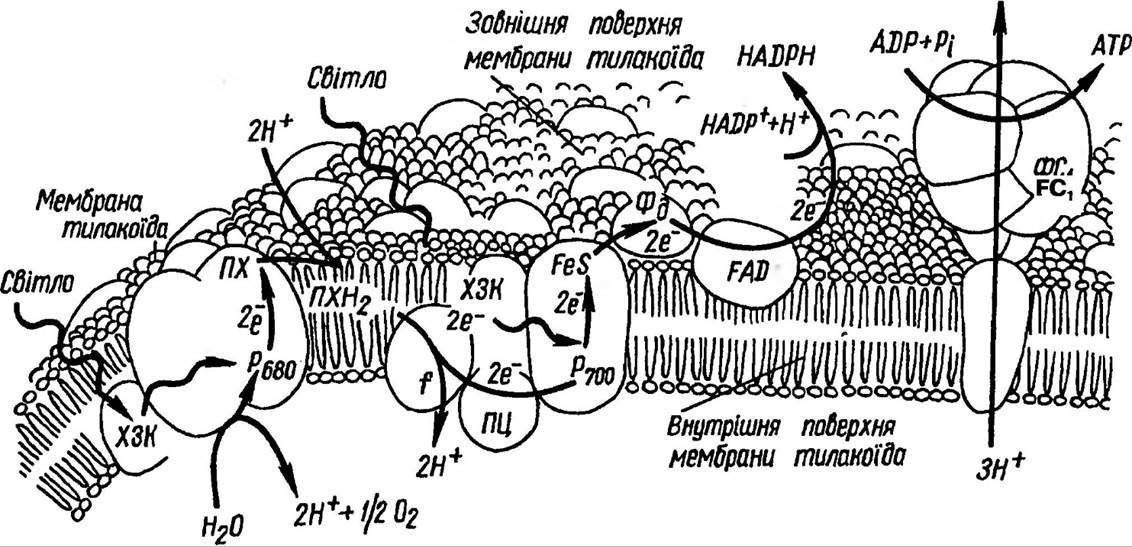
Рис 63. Схематичне зображення ланцюга перенесення електронів, яке пояснює, яким чином ця система транспортує через мембрану тилакоіду протони та здійснює синтез АТФ
Нециклічне транспортування електронів. Сонячна енергія, яка поглинається світлозбираючим комплексом фотосинтетичних одиниць, передається в реакційні центри до Р690 і Р700, що також здатні поглинати світло безпосередньо. Від фотозбуджених молекул хлорофілів реакційних центрів Р690 і Р700 відповідно, відриваються пари електронів, у результаті чого в молекулах залишаються ділянки з надлишковим позитивним зарядом Ф, які називаються «дірками». Електрони від Р690 (2е ) спочатку переносяться на феофітин, а далі уловлюються особливою формою переносника електронів — пластохіноном. У цій точці мембрани за електронами слідують додатково ще два протони із строми, яка розташована зовні від тилакоїду, у результаті чого пластохінон відновлюється до Пх×Н2 (рис. 64).
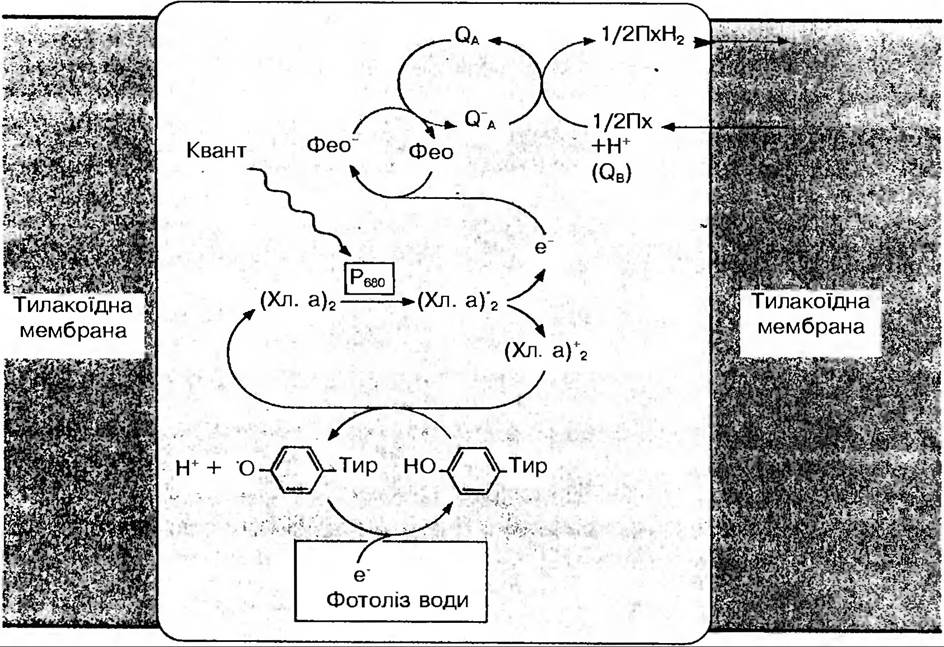
Рис. 64. Схема електронного транспорту у фотосистемі II
Протони потім звільняються і надходять всередину тилакоіду, а електрони передаються спочатку цитохрому f, потім пластоціаніну (ПЦ) і, нарешті, до Р700, де вони заповнюють "дірку».
Надалі електрони від Р700 уловлюються білком Fx, що містить залізо і сірку, після чого передаються фередоксину. Що таке окислювально-відновна система, яка стоїть першою у ланцюгу перенесення е від Р700 до НАДФ+ до цього часу не відомо. Тому її, як правило, позначають X. Систему виявлено за характерним сигналом електронного парамагнітного резонансу, який з’являється під час фотоокислення Р700. Побічна оцінка редокспотенціалу цієї системи дала велику від’ємну величину — 0,73 В. Отже, для того щоб відновити X до X-, Р700 повинен бути сильним відновником і мати потенціал більш від’ємний, ніж —0,73 В. Таким чином, X відновлює фередоксин.
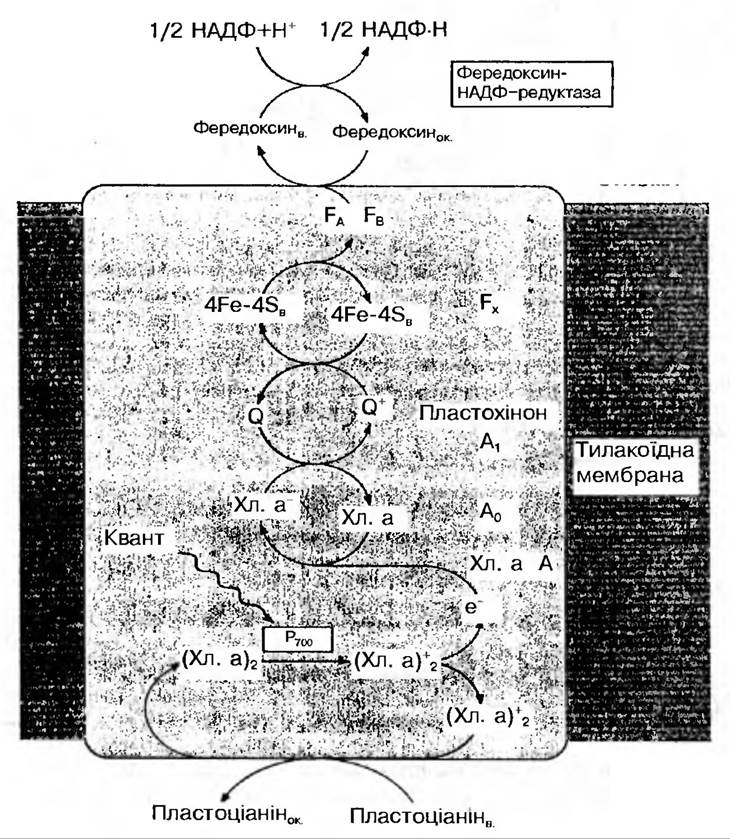
Рис 65. Схема транспортування електронів у фотосистемі І
Є дані, що свідчать про те, що в ході нециклічного перенесення електронів переносник X відновлює один із фередоксинів, пов’язаних з мембраною, можливо ФД з Eq — 0,54 В, а той, у свою чергу, відновлює розчинний фередоксин. (Вважають, що ФД з Е0 — 0,59 В бере участь у циклічному транспортуванні електронів.).
Розчинний фередоксин у хлоропластах утворює комплекс у співвідношенні 1 : 1 з такою окислювально-відновною системою, як фермент-фередоксин-НАДФ-оксиредуктаза, тобто ФАД-вмісним флавопротеїном. Вважають, що цей комплекс слабко зв’язаний із зовнішньою поверхнею мембрани тилакоїду. Для відновлення ФАД фередоксин-НАДФ-редуктази треба 2 електрони і 2Н+. Протони надходять із водного оточення (цим урівноважується виділення двох протонів з молекули води при фотоокисленні її), а електрони — від двох молекул відновленого розчинного фередоксину. Після цього відновлений флавопротеїн відновлює НАДФ+. Оскільки НАДФ — це двоелектронна окислювально-відновна система, то для утворення однієї молекули НАДФ×Н треба 2 електрони. Отже, після того як у пігментній системі ФС І відбудеться поглинання двох фотонів, передача поглинутої енергії і збудження молекули хлорофілу РЦ, пара електронів повинна бути перенесена по ланцюгу перенесення електронів від Р700 до НАДФ+ (рис. 66).
Віддаючи електрон акцептору X, фотозбуджений хлорофіл Р700 перетворюється на вільний катіон-радикал Р700 Для того щоб нециклічне транспортування електронів могло продовжуватися, треба передати електрон на Р700+, тим самим відновити його, повернувши у вихідний стан
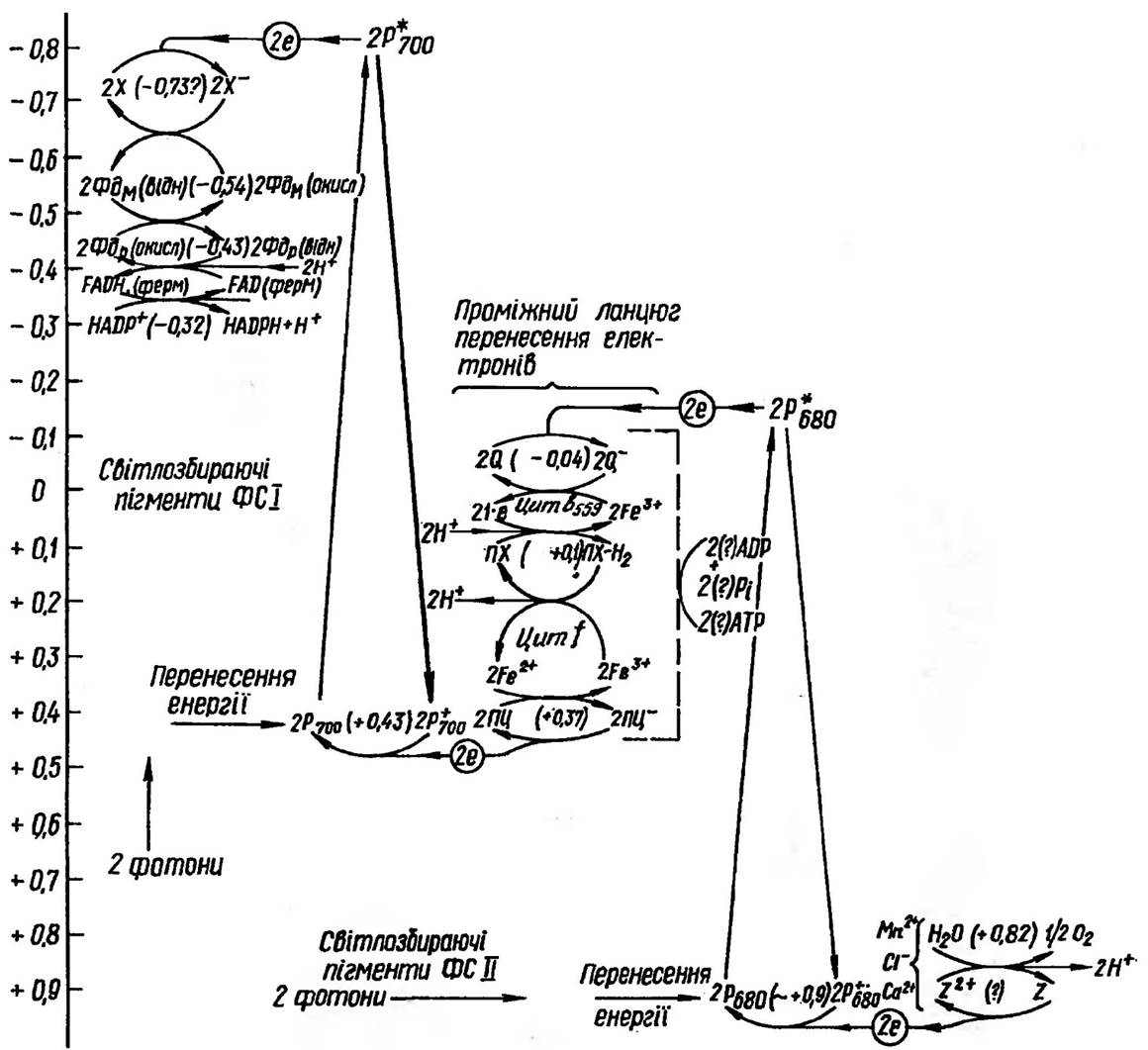
Рис. 66. Ланцюг окислювально-відновної системи на шляху від води до НАДФ+. Фдм — зв'язаний з мембраною фередоксин; Фдр — розчинний фередоксин; Q, Z, X — неідентифіковані окислювально-відновні системи
Р700. Система Р700/Р700+ — одноелектронна окислювально-відновна система (E0 = +0,43 В). Тому P700+ може бути відновлений пластоціаніном, більш електровід’ємною редокс-системою з Е0 = +0,37 В. Окислений Пц відновлюється цитохромом f. Як і решта цитохромів, це одноелектронна редокс-система (Ео = +0,36 В).
Окислений цитохром f в свою чергу відновлюється двоелектронною редокс-системою Пх/Пх×Н2 з потенціалом E0 + 0,1 В. Система Пх/Пх×Н2 переносить разом з двома електронами ще два протони. Ці протони мають важливе значення для синтезу АТФ. Коли Пх відновлюється двома електронами, одержаними від цитохрому b 559 (даний цитохром передує Пх в електрон- транспортному ланнцюгу), два протони переносяться із зовнішньої сторони тилакоїду. А коли Пх×Н2 відновлює дві молекули цитохрому f, два протони звільняються і потрапляють у внутрішній простір тилакоїду.
Отже, у міру того, як два е передаються по ЕТЛ через систему Пх/Пх×Н2, два протони переносяться через мембрану тилакоїду. Вважають, що градієнт концентрації протонів, який виникає при цьому (висока концентрація Н і низька ОН всередині тилакоїду і, навпаки, низька Н + і висока ОН зовні тилакоїду), і є рушійною силою процесу синтезу АТФ.
Як наслідок, спостерігається підвищення концентрації протонів усередині тилакоїдів і виникнення значного градієнта pH між зовнішньою і внутрішньою сторонами тилакоїдної мембрани: з внутрішньої сторони середовище виявляється більш кислим, ніж зовні. Підвищена кислотність всередині тилакоїду ще збільшується внаслідок фотоокислення води, причому електрони і О2 відходять (електрони заповнюють «дірку» у молекулі Р680, а О2 виділяється у вигляді газу), а протони накопичуються.
Особливо варто підкреслити, що в хлоропластах пластохінон міститься в таких кількостях (5-10% усього вмісту хлорофілу), що значно перевищує наявність усіх інших систем (наприклад, ПЦ —0,2%, цитохроми — 0,25% від загальної концентрації хлорофілів). Пластохінон, як вже було зазначено, відновлюється цитохромом b559, який існує в двох формах, що взаємоперетворюються, з різними значеннями Е0. Форма з низьким потенціалом, що відновлює пластохінон, має значення E0 приблизно +0,06 В. Форма з високим потенціалом, яка може відновлювати Р680, має Е0 + 0,37 В і не бере участі у нециклічному перенесенні електронів. Низько- потенціальний цитохром b559, в свою чергу отримує електрон від відновленого Q (неідентифікована система Q/Q- з потенціалом -0,04 В, яка править за первинний акцептор електронів від фотозбудженого Р680).
Таким чином, електрони в проміжному ланцюгу переносяться з рівня E0 -0,04 В на рівень +0,37 В. При цьому виділяється енергія, яка запасається у вигляді градієнта концентрації протонів. Вона використовується при зворотному пересуванні протонів зсередини назовні тилакоїдної мембрани особливими каналами АТФ-синтетази для синтезу АТФ. Протони йдуть по каналах доти, доки їхня концентрація всередині тилакоїду перевищить зовнішню, а отже, поки електрони під впливом поглиненого хлорофілом світла пересуваються по ЕТЛ.
Можна вважати, що роль ФС II полягає у тому, щоб підняти електрони від потенціалу води (E0 +0,82 В) до від’ємного потенціалу, який потрібний для виходу на початок проміжного ланцюга перенесення.
Після того як електрони проходять по ньому за градієнтом потенціалу і досягають кінця проміжного ланцюга та забезпечують синтез АТФ, їх знову треба підняти на рівень з досить від’ємним потенціалом, щоб вони могли відновити НАДФ+ (Е= -0,32 В). Саме цю роботу виконує ФС І.
Циклічне транспортування електронів. Шлях транспортування електрона у даному процесі точно не встановлено. Вважають, що він відбувається від X- на Фд з E0 = 0,59 В (один із пов’язаних з мембраною фередоксинів, про які йшлося раніше) і далі через цитохром b6 (Ео = -0,18 В) на пластохінон.Від відновленого Пх-Н2 електрони передаються на цитохром, далі на пластоціанін і на Р700. Оскільки до складу цього ланцюга перенесення електронів входить пластохінон, то перенесення електронів супроводжується синтезом АТФ, ймовірно 2АТФ/2е-. Циклічне транспортування включає, таким чином свого роду «обернену реакцію», в ході якої електрони через посередництво певних кофакторів і цитохромів, знову повертаються до хлорофілу Р700, що був окислений при первинній реакції. Тим самим вони заповнюють «дірку» (електронну вакансію), завдяки чому цей хлорофіл може знову поглинати фотони та переходити в фотозбуджений стан.
Можлива структурна організація мембрани тилакоїду, яка враховує як нециклічне, так і циклічне транспортування електронів навколо ФС І, запропонована Говінджі (рис. 67).
Фотосинтетичне фосфорилування (ФФ). Джерелом вільної енергії у всіх фототрофних організмів є енергія поглинених квантів світла. Для того, щоб Клітина могла використати цю енергію для біохімічної роботи, необхідно перетворити її в лабільну форму фосфатних зв’язків АТФ в процесі ФФ.
Фотосинтетичне фосфорилування, відкрите Д. Арноном в 1954 р., є спряжений з фотоіндукованим перенесенням електронів в хлоропластах процес синтезу АТФ. Обидва розглянуті потоки електронів поєднуються з синтезом АТФ, у зв’язку з чим розрізняють два основні типи фосфорилування: циклічне і нециклічне (табл. 6).
В даний час існують методичні прийоми експериментального розділення різних типів фосфорилування. Цього досягають шляхом додавання в раекційну суміш з ізольованими хлоропластами відповідних кофакторів, специфічних інгібіторів, вибірковим активуванням однієї або двох фотосистем світлом з різною довжиною хвиль, створенням аеробних чи анаеробних умов.
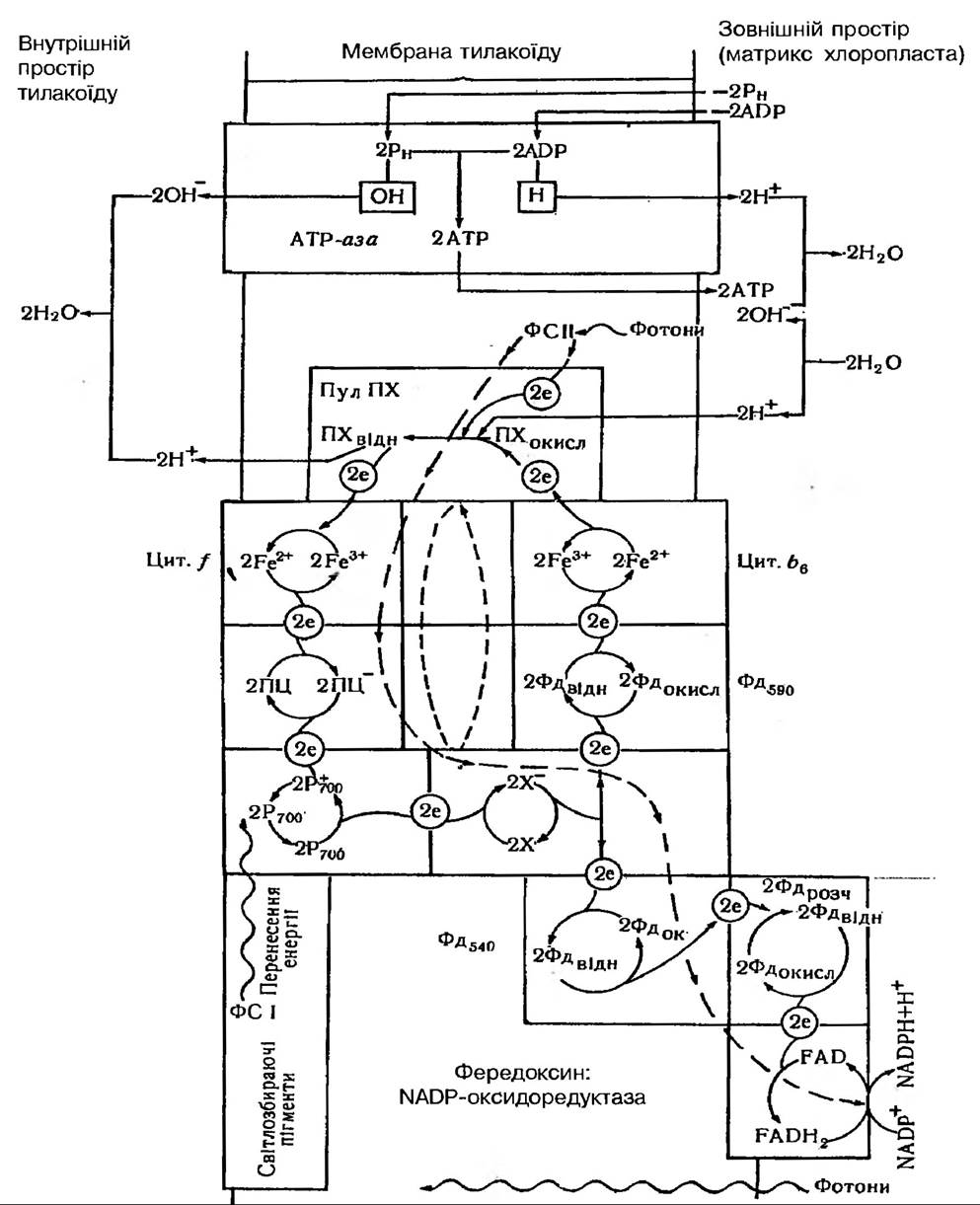
Рис. 67. Можливе розміщення окислювально-відновних систем в мембрані тилакоїда та ймовірна схема циклічного і нециклічного перенесення електронів, спряженого із синтезом АТФ
Таблиця 6. Порівняння циклічного та нециклічного фотофосфорилування
|
Відмінність процесів |
Нециклічне |
Циклічне |
|
Шлях електронів |
Нециклічний |
Циклічний |
|
Первинний донор електронів (джерело електронів) |
Фотосистема 1 (Р700) |
|
|
Останній акцептор електронів |
НАДФ* |
Фотосистема 1 (Р700) |
|
Кінцеві продукти |
АТФ, НАДФ×Н2, О2 |
АТФ |
|
Фотосистеми, які беруть участь у процесі |
ФС I та ФС II |
ФС І |
Циклічне ФФ поєднується з циклічним потоком електронів за участю лише ФС І. При фотозбудженні пігменту реакційного центру ФС І Р700 відбувається розподіл зарядів на мембрані. Енергія розділених зарядів може бути використана для синтезу АТФ в реакціях циклічного ФФ.
В такому випадку електрон через ряд проміжних переносників повертається знову до первинного донору електронів — Р700, тоді як енергія, звільнена в окислювально-відновних реакціях, використовується для синтезу АТФ.
Процес циклічного ФФ є найдревнішою формою фіксації енергії. Він не супроводжується ні виділенням О2, ні синтезом відновлених кофакторів і відображається рівнянням:
![]()
Шлях електронів у циклічному ФФ залежить від pH, редокс-стану переносників та медіаторів процесу, що визначає можливість замикання циклічного потоку електронів в різних ділянках ЕТЛ. Внаслідок замикання циклу від відновлених електровід’ємних компонентів ЕТЛ на електропозитивні переносники виникають різного типу альтернативні циклічні потоки. Від включення тих чи інших кофакторів залежить циклічний шлях, число спрягаючих центрів і загальний енергетичний вихід АТФ.
Нециклічне ФФ поєднується з потоком електронів від води до НАДФ+. Цей тип ФФ є еволюційно більш пізнішим і вимагає спільної дії обох фотосистем. Енергія світлових квантів, що поглинається пігментним комплексом ФС І та ФС II, перетворюється не лише в хімічну енергію макроергічних зв’язків АТФ, а і в хімічний потенціал відновленого НАДФ та витрачається на виділення О2 під час фотолізу води:
![]()
Нециклічне ФФ відбувається в аеробних умовах.
До нециклічного типу ФФ відноситься і псевдоциклічне ФФ. Акцептором електронів в даному випадку служить кисень. В результаті утворюється Н2О і загальна кількість кисню, що виділяється в ході фотолізу води, врівноважується кількістю відновленого О2, а тому видимого виділення кисню не спостерігається. Псевдоциклічне ФФ характеризується невисоким енергетичним виходом. Циклічне і нециклічне ФФ прийнято включати до світлової фази фотосинтезу, хоча світло необхідне лише на перших етапах цих процесів. ПЕРЕНЕСЕННЯ ЕЛЕКТРОНІВ В ЕТЛ може відбуватися і в темряві.
Як же здійснюється зв’язок між перенесенням електронів і утворенням АТФ за рахунок енергії, що виділяється при цьому? Найбільше визнання одержала хеміосмотична гіпотеза англійського вченого П. Мітчела (рис. 68). Як відомо, переносники електронів і протонів в
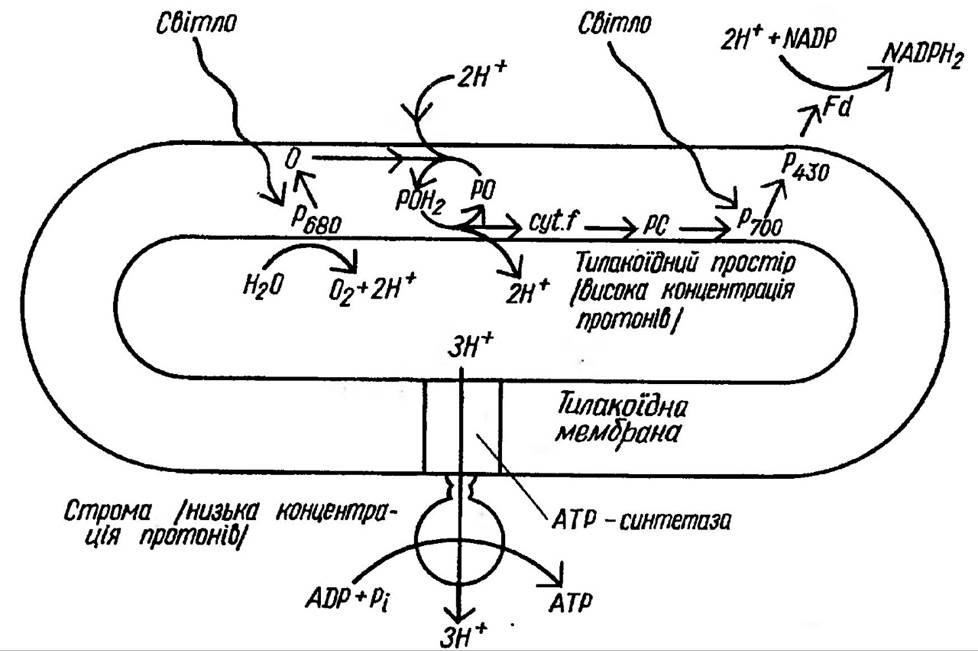
Рис. 68. Ілюстрація гіпотези хеміосмотичного спряження як механізму фотосинтетичного фосфорилування
тилакоїдній мембрані локалізовані асиметрично. Внаслідок активного перенесення протонів із строми у внутрішньотилакоїдний простір на мембрані створюється електрохімічний потенціал водню (ΔμН+), який складається із двох градієнтів: ΔрН — концентраційного, що обумовлений нерівномірним розподілом іонів Н+ по різні сторони мембрани, і електричного, обумовленого протилежним зарядом різних сторін мембрани (завдяки накопиченню протонів з внутрішньої сторони мембрани). Відповідно і енергія, накопичена на мембрані має дві складові: концентраційну та електричну. Щоб використати цю енергію, необхідно забезпечити зворотний потік
протонів. Енергія, яка виділяється при цьому, використовується для синтезу АТФ. При перенесенні 2е від первинного донора (+0,82 В) до кінцевого акцептора (-0,32 В) за участю двох систем величина окислювально-відновного потенціалу складає -1,14 В, що еквівалентно 220 кДж (П.Г. Костюк та ін., 1988). Відомо, що вільна енергія одного макроергічного зв’язку дорівнює 34,54 кДж/моль. Значить, для синтезу однієї молекули АТФ із АДФ і неорганічного фосфату необхідна саме така кількість енергії. В ряду переносників цій умови відповідають донорно-акцепторні пари:
пластохінон (0,36 В) і цитохром f (0,11 В)
ΔЕ0 = 0,25 В
Цитохром b559 (0,06 В) і Р700 (0,43 В)
ΔЕо = 0,37 В
Н2О <=> 1/2О2 + Н2 (0,82 В) і Р680 (-0,05 В)
ΔЕ0 = 0,87 В.
З цими ділянками ЕТЛ і пов’язують пункти спряження ФФ. Існують різні гіпотези відносно механізму фосфорилування. Допускають, що фосфатна група та АДФ можуть безпосередньо зв’язуватися з ферментом АТФ-синтетазою в активній ділянці комплексу CF, (див. рис. 59). Тоді два протони переміщуються через канал CF0 за градієнтом концентрації і, поєднуючись з киснем фосфата, утворюють воду. Це робить фосфатну групу досить реактивною, і вона приєднується до АДФ з утворенням АТФ.
Згідно інших поглядів, АДФ і Фн поєднуються в активному центрі фермента спонтанно. Однак, утворена АТФ міцно зв’язується з ферментом, і для її звільнення необхідна енергія. Цю енергію постачають протони, які, зв’язуючись з ферментом, змінюють його конформацію, після чого АТФ звільнюється (рис. 69).
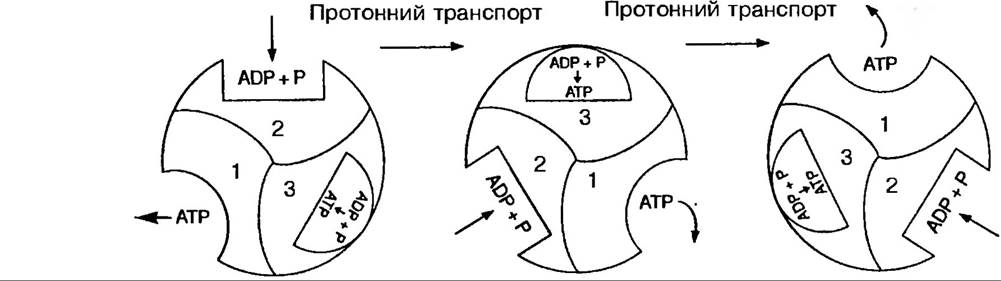
Рис. 69. Ймовірний Механізм синтезу АТФ
Продукти світлової фази, в яких акумульована енергія сонячних квантів, АТФ і НАДФ×Н2, Д. Арнон назвав асиміляційною силою. їх утворення відбувається за відсутності СО2.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.