Фізіологія рослин - Мусієнко М.М. 2001
Фізіологія рослин та біотехнологія. Досягнення і перспективи розвитку
Біологічна фіксація азоту та генетична інженерія
На долю промислового синтезу аміаку в світі припадає 1/4 частина азоту, тоді як 3/4 припадає на біологічний синтез. За останні роки повернення азоту в грунт в Україні все ще не перевищує 75%, а під зерновими — 60-63% від винесеного з урожаєм. Ліквідація азотного дефіциту, особливо в останнє десятиріччя XX століття, пов’язана із значними труднощами енергетичного, екологічного та економічного порядку.
Є дані, що внесення значних кількостей мінерального азоту (150-300 кг/га) пов’язане із токсичністю іонів NO3-, NH4+. Зокрема, в таких умовах в рослинному організмі збільшується вміст небілкового азоту, непротеїногенних амінокислот, змінюється гормональний статус рослин, знижується відтік асимілятів, проникність мембран. Все це впливає на Метаболізм азоту і вуглеводів, а в кінцевому етапі на кількісний та якісний склад продуктів фотосинтезу. Саме тому в багатьох країнах Західної Європи, США існуючу систему землеробства намагаються замінити більш раціональною біодинамічною системою.
При вирішенні азотної проблеми біологічним шляхом найбільш доцільним є використання бобово-ризобіальних систем, як найбільш економічно вигідних (вони накопичують від 40 до 300 кг азоту в рік на 1 га). Вивчення генетики бульбочкових бактерій та молекулярно-генетичні дослідження генів азотфіксації дозволяють створити умови для цілеспрямованого конструювання господарсько цінних штамів ризобій, а в перспективі перейти до створення нових азотфіксуючих симбіозів з різними культурами.
Найбільш дійовим і ефективним шляхом підвищення азотфіксації симбіотичними системами бобових є застосування бактеріального препарату ризотрофіну (нітрагіну).
Приріст урожаю від застосування нітрагіну становить 10-15%, а для культур, нових для даного регіону, які не мають спонтанних бульбочкових бактерій, — і до 100%. Позитивна дія нітрагіну не обмежується цим, адже він значно поліпшує якість продукції, зокрема, вміст білків та жирів.
Основне завдання щодо підвищення ефективності нітрагінізації — одержання високоактивних, вірулентних штамів, стійких до антагоністів, здатних конкурувати з мікрофлорою грунту, зокрема, малоактивними бульбочковими бактеріями.
В останні роки доведено, що основні симбіотичні властивості в більшості бактерій локалізовані на великій плазміді sym з молекулярною масою 85-260 мегадальтон. Це відкриває можливості для поліпшення господарсько цінних ознак у бульбочкових бактерій шляхом перенесення плазмід. Прогрес молекулярно-генетичних досліджень у ризобій створив умови, які дозволяють маніпулювати симбіотичними генами методами генетичної інженерії.
Симбіотична фіксація азоту забезпечується з боку бульбочкових бактерій генами симбіозу (nod- або sym-гени), які контролюють специфічність бактерій до певного виду бобових рослин, зараження кореня та утворення бульбочок. Крім того, вони мають власне гени азотфіксації (nif- або fix-гени), які кодують комплекс ферментів на чолі з нітрогеназою. І, нарешті, третя група генів, яка бере непряму участь в азотфіксації (регуляторна функція, забезпечення енергією і т. д.).
У найближчій перспективі буде виконано роботи щодо направленого конструювання перспективних штамів бульбочкових бактерій. Вже сконструйовано рекомбінантну р-плазміду pRD1, яка несе ген із Klebsiella pneumoniae. її використано для надання азотфіксуючої здатності 14 видам мікроорганізмів, і у восьми з них одержано експресію генів азотфіксації. Успішні роботи щодо перенесення генів азотфіксації (nif -генів) з Е. соїі і експресія їх в новому хазяїні продемонструвала можливість міжродового перенесення генів азотфіксації (рис. 214).
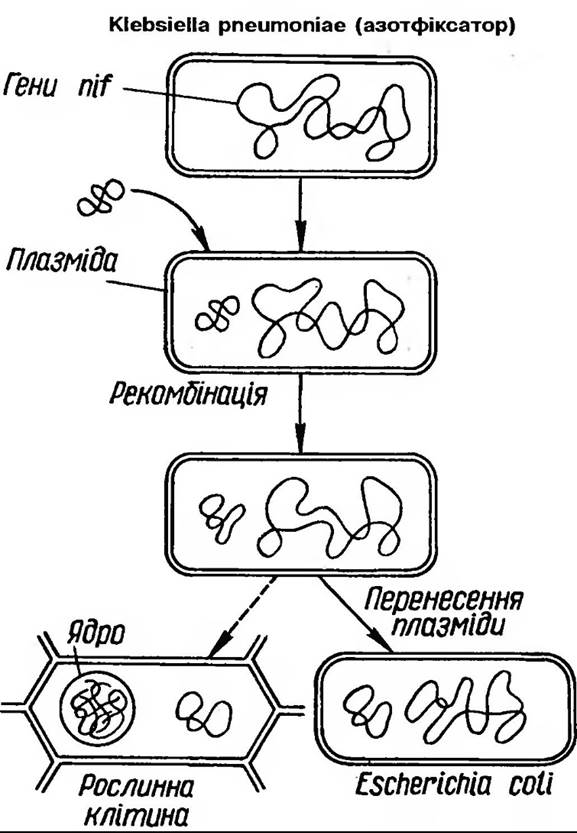
Рис 214. Перенесення генів, які відповідають за азотфіксацію, в рослинну клітину
В Інституті фізіології рослин та генетики НАН України в лабораторії Ю.П. Старченкова в результаті міжродової кон’югації Е. соlі, яка містить плазміду pRD1, та бульбочкових бактерій одержано більше 70 транскон’югантів ризобій конюшини, люцерни, машу. Виявлено, що вони характеризуються різною стабільністю за стійкістю до антибіотиків, а також відрізняються від вихідного штаму та між собою за деякими фізіолого-біохімічними та симбіотичними властивостями. Ці результати свідчать про гетерогенність одержаних транскон’югатів і можливість використання транскон’югатів з підвищеною азотфіксуючою активністю та ефективністю як вихідного матеріалу для селекції виробничих штамів бактерій.
Конструювання і відбирання перспективних штамів бульбочкових бактерій насамперед повинні забезпечувати підвищення енергетичного коефіцієнта корисної дії симбіотичної азотфіксації. Справа в тому, що в процесі фіксації азоту нітрогеназний комплекс поряд з відновленням N2 каталізує також і АТФ-залежне відновлення протонів з утворенням Н2. Виявлено, що кількість водню, яка виділяється, залежить від його рециклізації, тобто від наявності в бактероїдах Н2-зв’язуючої гідрогенези, яка б забезпечувала реутилізацію виділюваного нітроге-
назою водню. У зв’язку з тим що процес зв’язування водню зумовлює утворення АТФ, він відіграє істотну роль в енергетиці азотфіксації. В результаті такі симбіотичні системи зв’язують на 14-31% молекулярного азоту менше, ніж ті, що не виділяють водень, тобто в бактероїдах яких функціонує Н2-зв’язуюча гідрогеназа. Вона, як виявилось, є далеко не у всіх видів і штамів бульбочкових бактерій.
Зараз інтенсивно ведуться роботи щодо вивчення біохімічних та генетичних особливостей взаємозв’язку між нітрогеназною та Н2-окислюючою гідрогеназною активністю з метою з’ясувати умови, при яких Н2-зв’язуюча гідрогеназа бобових була б особливо активною.
Деякі штами бульбочкових мають так звані Hyp-гени (від англ, hydrogen uptake — поглинання водню), що кодують фермент гідрогеназу. Тому одним з основних моментів при конструюванні перспективних штамів бактерій повинно стати включення в Геном бактерій Нур-генів.
ГЕНЕТИЧНА ІНЖЕНЕРІЯ РОСЛИН та азотфіксація. Можна припустити, що перенесення угруповань генів більш простих, ніж ціла п/Т-область, цілком імовірне. В ролі векторів можуть бути використані віруси рослин, наприклад вірус мозаїки кольорової капусти — дезоксирибовірус, до ДНК якого можна приєднати певні nif-гени, які можуть бути інтегровані в геном рослин або транскрибуватися в рослинній клітині одночасно з ДНК-вірусом при його реплікації. Можливий і інший вектор, наприклад патогенна бактерія Agrobacterium tumefaciens, яка містить крупну плазміду Ті, що відповідає за індукцію пухлин в рослинних тканинах (корончатий гал).
Особливий фрагмент Ті-плазміди, названий т-ДНК (від англ, transferred DNA — ДНК, яка переноситься), може інтегруватися в геном рослин. т-ДНК має молекулярну масу (10-15) × 106 дальтон і складається щонайменше з трьох функціональних одиниць. Нуклеотидні послідовності розташовані на кінцях тДНК досить подібні із послідовностями певних ділянок нетрансформованої клітинної ДНК, а тому рекомбінація між цими двома ДНК зумовлює включення тДНК в геном рослин та переродження інфікованих тканин.
Отже, утворення пухлин в тканинах рослин, що спричинюється Ті-плазмідою Agrobacterium tumefaciens, індукується перенесенням сегмента бактеріальної ДНК (із прокаріотної клітини) та інтеграцією його в геном евкаріотних клітин. З таких пухлин вдається регенерувати повноцінні рослини, які містять в своїх клітинах тДНК. До складу тДНК аналогічно можна перенести спеціальні гени та одержати рослини, які будуть містити в своїх клітинах ці гени.
Для підвищення вкладу біологічної азотфіксації у виробництво, великий інтерес має Введення в клітину небобових рослин генів, які забезпечують азотфіксуючий симбіоз з бульбочковими бактеріями у бобових рослин. Можливо, це буде один з небагатьох доступних шляхів для створення нових азотфіксуючих симбіозів і система тДНК агробактерій відіграє тут істотну роль.
Слід, однак, зазначити, що оскільки Agrobacterium tumefaciens інфікує лише дводольні рослини, то буде не простою справою введення Ті-плазміди для перенесення специфічних генів в клітини однодольних рослин, зокрема хлібні злаки.
До того ж при використанні в ролі вектора агробактерій виникають нові проблеми, пов’язані з регуляцією експресії цих nif-генів, зокрема, як захистити нітрогеназу від дії О2. Та, незважаючи на все це, Методи планування та рекомбінації нуклеїнових кислот створили передумови для перенесення генів в клітини рослин та експресію їх. Гени nif можна перенести і в ендотрофну мікоризу, так як перенесення в клітини гриба здійснити навіть легше.
Однак незалежно від методів, якими перенесені nif-гени, експресія, тобто Транскрипція їх в мРНК та трансляція останніх в білок, повинна здійснюватися в рослинній клітині. У разі локалізації в хромосомі для експресії nif-генів необхідне злиття його з евкаріотичним промотором (у разі агробактерій для цієї мети може бути корисним промотор області тДНК Ті-плазміди). Адже механізми експресії генів в хлоропластах та в клітинах прокаріот досить близькі.
Слід пам’ятати, що здатність до азотфіксації залежить не лише від наявності nif-генів, а і генів, які відповідають за синтез переносників електронів та факторів, які потрібні для нормального функціонування нітрогенази. Саме тому створення рослин, здатних фіксувати азот незалежно від симбіотичних мікроорганізмів, залишається досить складним завданням. Скоріше можна досягти успіхів у вивченні факторів, які індукують симбіоз, щоб підвищити його ефективність.
Дослідження в галузі фізіологічної генетики рослин внесуть вагомий вклад у відбір більш ефективних рослин-господарів. Соматична гібридизація, можливо, дозволить одержати нові сорти рослин, які будуть здатні до симбіотичної азотфіксації. Цілком імовірно, що буде модифіковано і геном мікроорганізмів, який утворює ризосферу із небобовими рослинами з метою підвищення ефективності таких асоціацій.
Тому фундаментальні дослідження щодо перенесення генів азотфіксації в клітини вищих рослин та їх експресії мають багатообіцяючу перспективу. Одержані результати вже досягли рівня, коли стало можливим конструювання перспективних штамів методами молекулярної біології та генної інженерії. Створено умови для генної інженерії рослин на основі онкогенних плазмід бактерій. Одержано перші результати, які продемонстрували можливість та доцільність поєднання досліджень генної інженерії азотфіксації та генної інженерії рослин.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.