Фізіологія рослин - Мусієнко М.М. 2001
Фізіологія виділення речовин рослинним організмом
Мехашзми виділення речовин
Виділення речовин у рослин може здійснюватись за участю різноманітних механізмів, часто проти концентраційного градієнта із затратами енергії АТФ. Розрізняють мерокринову (meros — частина, crino — виділяю), апокринову та голокринову секреції.
Мерокринову секрецію поділяють на:
мономолекулярну (еккринову) секрецію, яка здійснюється з допомогою іонних насосів через мембрани;
гранулокринову, де виділення речовин, в певних мембранних упаковках (везикулах, секреторних пухирцях оточених мембранами тощо), здійснюється всередину клітини чи певні компартменти її;
секрецію із цистерн ендоплазматичної сітки безпосередньо на поверхню плазмалеми.
Апокринова секреція відбувається шляхом відокремлення разом із секретом або частини цитоплазми, або, наприклад, голівок у сольових волосків деяких галофітів. Використовується також під час секреції ліпідів.
Голокринова секреція характеризується тим, що при цьому відбувається перетворення всієї клітини в секрет, наприклад виділення слизу клітинами кореневого чохлика.
На рівні кожної клітини секреторні процеси здійснюються завдяки мономолекулярній та гра- нулокриновій секреції. Активний транспорт в мембранах рослинних клітин здійснюють протонні Н+ —АТФ-ази, Na+, К+, Са2+-АТФ-ази, аніонні АТФ-ази тощо. Іонні насоси, які забезпечують мономолекулярну секрецію, можуть функціонувати в плазмалемі, тонопласті, інших мембранах органел клітини.
Секреторний процес на рівні клітини включає поглинання та накопичення вихідних продуктів, синтез поліцукрів, білків, терпеноїдів та концентрування їх у вигляді секрету, нарешті виділення секрету з наступним відновленням структур клітини. На клітинному рівні секрецію можуть здійснювати також спеціалізовані секреторні ідіобласти.
Провідну роль в гранулокриновій секреції відіграє Апарат гольджі, в диктіосомах якого і концентруються компоненти майбутнього секрету. Вважають, що сполуки, які проникають в диктіосоми із гранулярного ендоплазматичного ретикулуму, де вони синтезуються, спочатку заключаються в мембрану і вже потім у везикулярній формі переносяться до плазмалеми. В процесі злиття везикул з плазмалемою вміст їх виявляється між плазматичною мембраною та клітинною стінкою, тобто спостерігається екзоцитоз. Мембрана везикул монтується в плазмалему, забезпечуючи її оновлення та ріст, а компоненти секрету надходять до клітинної оболонки, вбудовуючись в її структури.
Встановлено необхідність іонів кальцію для здійснення секреції даного типу. Припускають, що кальцій знімає електростатичний бар’єр, який виникає між везикулою з секретом та плазмалемою, а також спричинює зміни мікротрубочок та мікрофіламентів, що направляють рух везикул до плазмалеми. Це відбувається при взаємодії іонів кальцію з білком кальмодуліном, який має чотири ділянки для його зв’язування. Утворення комплексу Сакальмодулін зумовлює скорочення мікрофіламентів та збирання — розбирання мікротрубочок цитоскелета, що сприяє переміщенню везикул по клітині (Т.С. Саламатова, 1991):
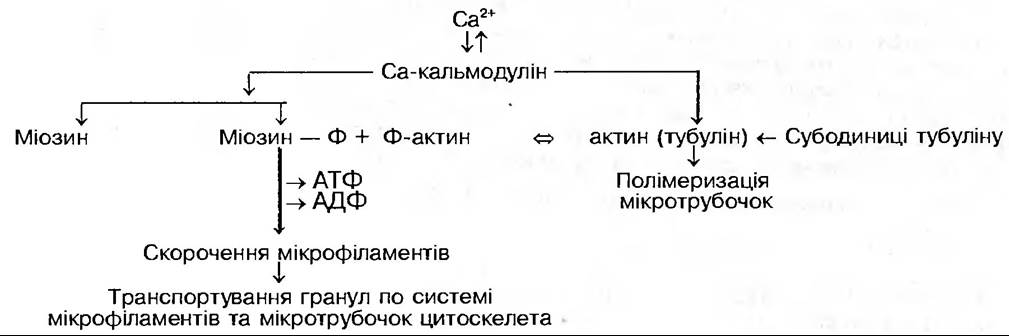
Кальцій знижує гідратаційний бар’єр при злитті мембран, знижує від’ємний заряд контактуючих мембран, створює кальцієві місточки між мембранами, забезпечує Са2+-залежне фосфорилування мембранних білків за участю кальмодуліну.
Гранулокринова секреція досить активно відбувається при верхівковому рості кореневих волосків, пилкових трубок, при формуванні фрагмопласту первинної стінки клітин, що діляться. Молекулярний механізм везикулярної секреції рослинних клітин потребує подальшого вивчення. Сучасні уявлення про їхні механізми дають змогу припустити спорідненість їх із аналогічними в тваринних організмах.
Системи виділення речовин у вакуоль
Вакуоль — внутрішній компартмент рослинної клітини, куди в процесі обміну речовин виводяться різноманітні речовини, частина яких можуть знову реутилізуватися. Тонопласт, тобто мембрана, яка оточує вакуолю має невисокий електричний опір (300-700 Ом×см2) і низький біоелектричний потенціал (20-40 мВ). Градієнт pH між вмістом вакуолей і гіалоплазмою (1-2 одиниці), більш висока концентрація речовин у ваколях в порівнянні з цитоплазмою вказує на існування активного транспорту іонів водню із цитоплазми через тонопласт у вакуоль, а також інших систем транспорту речовин. І дійсно, в тонопласті функціонує АТФ-залежна Н+ — помпа, яка забезпечує максимальний потік іонів Н+ у вакуоль (приблизно 15 нмоль/ см2×секунду).
Спрямований у вакуоль потік протонів створює електричний градієнт для транспорту сюди ж аніонів органічних кислот. Іони К+ можуть надходити у вакуоль в обмін на пасивний вихід із неї іонів водню. Надходження цукрів здійснюється за рахунок котранспорту поряд з потоком Н+. Таніни синтезуються в системі ендоплазматичного ретикулуму і попадають у вакуолю безпосередньо із його цистерн. Різні сполуки типу антоціанів перед транспортуванням у вакуолю перетворюються у розчинні глюкозиди і в такій формі переносяться у вакуоль.
У рослин, як відомо, запасні білки (розчинні альбуміни, досить щільний білковий матеріал з включенням фітину — кальцій-магнієвої солі інозитфосфорної кислоти, глобуліни тощо) відкладаються в клітині якраз у вакуолях. Існують різні механізми транспорту запасних білків, серед яких можливий шлях за допомогою переносників, попадання у вакуоль за рахунок інвагінації тонопласту, прямий вихід білків у вакуоль безпосередньо із цистерн гранулярного ендоплазматичного ретикулуму та інші (Саламатова, Зауралов, 1991).
Секреція лігніну, кутину та восків
У сучасних вищих рослин лігнін інкрустує стінки судин протоксилеми, а пізніше первинну та вторинну стінки всієї клітини. Процес лігніфікації відбувається впродовж всього життєвого циклу рослинних клітин, причому особливо інтенсивно після припинення їх росту. Вважають, що полімеризація лігніну відбувається між фібрилами целюлози безпосередньо в клітинній оболонці, хоча ароматичні спирти — основні мономери лігніну — синтезуються в цитоплазмі, наприклад, через пряме дезамінування ароматичної амінокислоти фенілаланіну. Лігнін вважається полімолекулою, в якій мономери зв’язані різними типами зв’язків: ефірними, вугле- вод-вуглеводними та дифенільними. Мономери постачаються в клітинну стінку шляхом дифузії (за градієнтом концентрації), або в мембранній упаковці.
Клітини епідермісу всіх органів рослин володіють здатністю до секреції компонентів кутину, які входять до складу кутикули. Кутин — гідрофобна полімерна речовина, яка складається із поліестерифікованих довгих молекул жирних кислот (С16-С32) з великим числом замісників. Попередники кутину, ймовірно, переносяться за допомогою гранулокриновоі секреції або дифузії через плазмалему і клітинну стінку на поверхню епідермальної клітини, де і перетворюються в кутин в процесі окислювальної полімеризації. При цьому шар кутину пронизується полісахаридними компонентами стінки (наприклад, целюлозою) і утворює кутикулу.
Поверхня кутикули багатьох рослин має шар ще більш гідрофобного полімера — воску. Воски — складні ефіри жирних кислот і високомолекулярних одноатомних спиртів. Кожен тип воску відкладається у вигляді точно визначених структур (їх близько 14 000) — від аморфної плівки до волокон і трубок різного розміру та форми. Вважають, що компоненти воску переносяться через клітинну стінку в « обгортці» із ліпо- або глікопротеїнів, які володіють зовнішніми гідрофільними поверхнями і тому можуть переноситись в гідрофільній клітинній стінці.
Спеціалізовані секреторні структури у рослин та функціонування їх. Структури видільної системи рослин дуже відрізняються як за ступенем спеціалізації, так і за локалізацією їх в рослинному організмі. Насамперед відзначимо відмінності секреторних структур відносно взаємозв’язку їх між секретом та протопластом секретуючої клітини. Виділюваний секрет може залишатись у видільній клітині або секретуватись назовні, в інші компартменти. Наприклад, Ефірні олії, бальзами, смоли, хоча і виступають як екскреторні речовини, досить часто і надовго накопичуються в клітині. Вони можуть розподілятися по цитоплазмі у вигляді вкраплень, можуть виділятися в міжклітинники або на поверхню рослини.
Секреторні структури можуть розміщуватись зовні, інші всередині певних органів рослини. Одні з них за структурою досить прості, у вигляді залозистих волосків, інші — багатоклітинні залози з власною провідною системою, а деякі мають вигляд міжклітинних ходів, порожнин.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.