Фізіологія рослин - Мусієнко М.М. 2001
Фотосинтез: фізіолого-біохімічні та екологічні аспекти
С4-шлях фотосинтезу
Цикл Кальвіна функціонує у вищих рослин, водоростей, фототрофних бактерій, але це не єдиний шлях фіксації вуглекислого газу. Як правило, інші шляхи є додатковими, тобто вони полегшують роботу ВПФ-циклу (рис. 72).
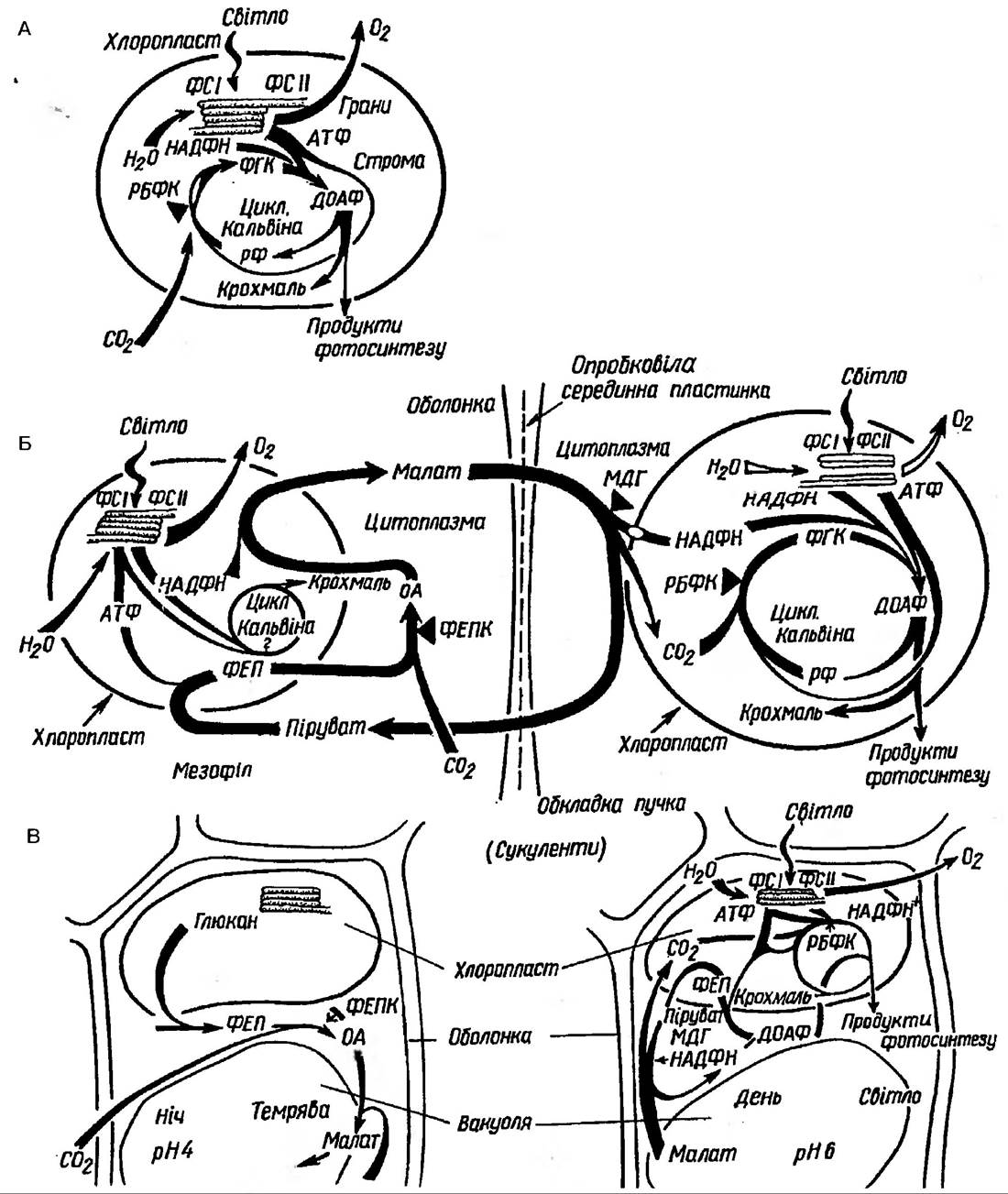
Рис 72. Шлях відновлення вуглецю в С3-рослин (А), С4-рослин із групи НАДФ×МДГ (Б) та CAM-рослинах з метаболізмом за типом Crassulaceae (В)
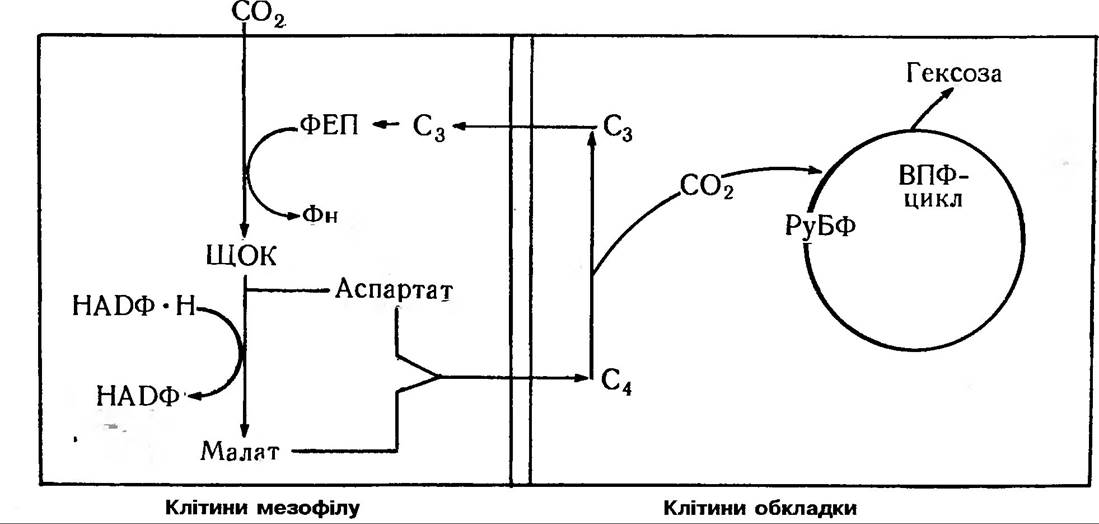
Рис 73. С4-шлях фотосинтезу (цикл Хетча і Слека)
Так, М. Хетч та К. Слек (1966) виявили, що в цукрової тростини та деяких інших тропічних рослин С14 в першу чергу з’являється в певних 4-вуглецевих сполуках (щавлевооцтова, яблучна, аспарагінова кислоти), тобто С4-дикарбонових кислотах. Такі рослини стали називати С4- типом рослин, на відміну від С3-типу рослин, у яких первинним продуктом темпової фази фотосинтезу є тривуглецеві сполуки — фосфогліцеринова кислота. Як виявилося, первинним акцептором СO2 у них є фосфоенолпіруват (ФЕП), а фермент карбоксилування — фосфоенолпіру- ваткарбоксилаза, тобто ФЕП-карбоксилаза (рис. 73).
Листки багатьох рослин С4-типу відрізняються від С3-типу своєю анатомічною будовою. Кожний провідний пучок в них оточений шаром досить великих паренхімних клітин — це так звана обкладинка судинного пучка, яка, в свою чергу, оточена дрібнішими клітинами мезофілу. Такий тип будови має назву «кранц-анатомії» — корончастий тип. Причому, хлоропласти в різних типах клітин, як виявилося, морфологічно різні: в клітинах обкладки — часто без гран, а в клітинах мезофілу добре виражені грани, але мало накопичується крохмалю. Було доведено, що в клітинах мезофілу вища активність ФЕП-карбоксилази, тоді як клітини обкладинки характеризуються високою активністю РуБФ-карбоксилази та інших ферментів циклу Кальвіна.
Отже, обидві групи клітин в С4-типу рослин кооперативно перетворюють СО2 на гексозу: в мезофілі ФЕП-карбоксилаза забезпечує приєднання СО2 до фосфоенолпірувату з утворенням відповідної дикарбонової кислоти. Далі вони дифундують в клітини обкладки і там декарбокси- луються з утворенням СО2 та піровиноградної кислоти (рис. 74).
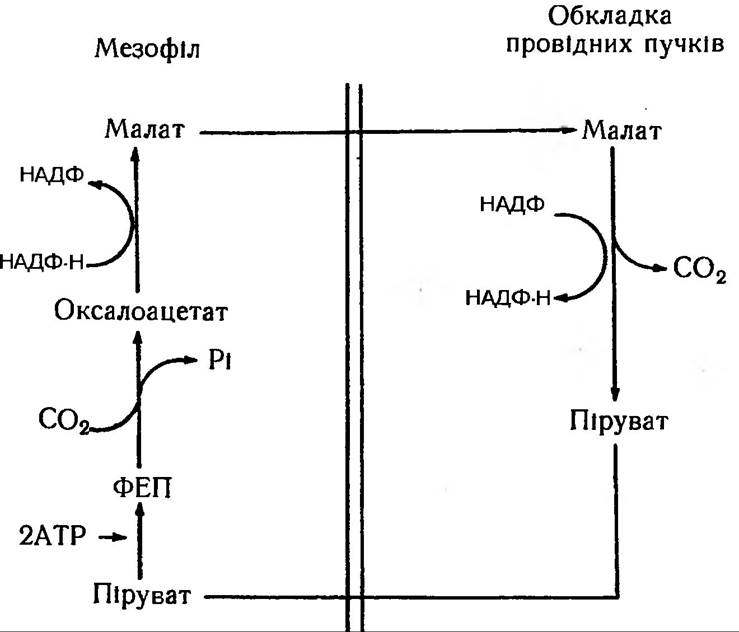
Рис. 74. С4-шлях фотосинтезу за участю НАДФ-малатдегідрогенази як декарбоксилуючого механізму
Піруват знову дифундує в мезофіл, де відбувається регенерація ФЕП, після чого карбоксилування повторюється за участю нової молекули СО2. Одночасно СО2, яка звільняється в клітинах обкладинки, надходить в цикл Кальвіна, тобто реагує з РуБФ, що спричинює утворення ФГК та інших продуктів, властивих С3-типу рослинам.
Отже, С4-типфіксації виступає чимось подібним до насоса, який постачає вуглекислий газ для С3-шляху фотосинтезу.
Причини появи С4-шляху в процесі еволюції недостатньо з’ясовані. Вони ідентифіковані лише серед наземних покритонасінних. Так, із приблизно 300 родин квіткових рослин, лише 16 включають в себе рослини С4-типу. Сюди відносять злаки, осоки та 14 родин дводольних. Більшість злісних бур’янів на Землі — рослини С4-типу,тоді як більшість С3-рослин — культурні рослини (табл. 7). Є також рослини з проміжним типом С3/С4. Так, у деяких видів проса (рід Panicum), немає чіткої диференціації між компартментами з ФЕП-карбоксилазою та РуБФ-кар- боксилазою, як це спостерігається у С4 рослин.
Таблиця 7. С3- і С4-типи рослин
|
Рослини |
С3 |
c4 |
|
Однодольні Дводольні |
Avena sativa L. Carex pendula L. Hordeum vulgare L. Iris sibirica L. Oryza sativa L. Phragmites communis L. Secale cereale L. Triticum aestuvum L. Cyperus papurrus L. Panicum lindheimevi Nash. Beta vulgaris L. Helianthus annuus L. Nicotiana tabacum L. Phaseolus vulgaris L. Atriplex hastata L. Euphorbia corollata L. |
Andropogon virginius L. Digitaria sanqiunalis (L.) Scop. Echinochloa coIonum L. Miscanthus sinensis L. Saccharum officinarum L. Sataria italica (L.) Beauv. Sorghum bicolor (L.) Moench. Zea mays L. Cyperus rotundus L. Panicum muliaceum L. Amatanthus albus L. Portulaca oleracea L. Salsora kali L. Kochia scoparis (L.) Roth. Atriplex rosea L. Euphorbia maculata L. |
Слід зазначити, що механізм С4-типу фіксації СO2 у різних груп рослин не однаковий. Залежно від того, як відбувається декарбоксилування С4-дикарбонових кислот у клітинах обкладки судинних пучків, усі рослини розподіляють на 3 групи (табл. 8).
Таблиця 8. Особливості декарбоксилування в різних груп рослин з С4-типом фотосинтезу
|
Тип |
Декарбоксилаза клітин обкладки провідних пучків |
Кофактори, які утворюються (+) або використовуються (-) під час декарбоксилування |
Основний субстрат, який транспортується |
|
|
з мезофілу у клітини обкладки |
з обкладки у мезофіл |
|||
|
I |
НАДФ +-малатдегідрогеназа |
+1 НАДФ×Н |
Малат |
Піруват |
|
II |
НАД- малатдегідрогеназа |
+ 1 НАД×Н |
Аспартат |
Аланін/піруват |
|
III |
ФЕП-карбоксикіназа |
-1 АТФ |
Аспартат |
Фосфоенол піруват |
Представники усіх трьох груп відрізняються також за локалізацією зазначених реакцій: власне в хлоропластах клітин обкладки (І група), у цитоплазмі і (або) мітохондріях (II і III група).
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.