Фізіологія рослин - Мусієнко М.М. 2001
Рухи рослин
Настії
Настичні рухи властиві тим органам, які мають двобічну симетричну будову. При цьому спостерігається нерівномірний ріст нижньої та верхньої частини органа. Якщо швидкий ріст спостерігається на морфологічно верхньому боці органа, то тут має місце епінастія, внаслідок чого розкриваються бруньки, бутони. В протилежному випадку, коли спостерігається швидкий ріст на нижньому боці, закриваються покриви квітки — явище гіпонастії. В більшості ж випадків настичні рухи обумовлені зміною тургору і проходять в результаті збільшення або зменшення концентрації осмотично активних речовин (малату, іонів калію, хлору) в спеціалізованих клітинах протилежних боків органа.
Можна спостерігати виникнення рухів, коли світло періодично змінюється темрявою. У деяких рослин квітки розкриваються вранці, а на ніч закриваються. Аналогічна властивість характерна і для листків деяких рослин (рис. 206).
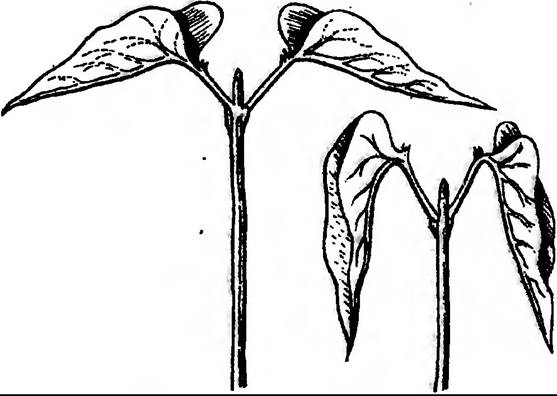
Рис 206. Ритмічні добові рухи положення листків Phaseolus multiflorus вдень (ліворуч) і вночі (праворуч)
Ці рухи дістали назву «сон рослин» — ніктинастії, хоч це й не відповідає названому фізіологічному стану організму і не має нічого спільного з явищем сну, властивого тваринам. «Сонні рухи» квітів та листків, коли вони відкриваються або закриваються, у відповідь на зміну світла (фотонастії) чи температури (термонастії) належать до настичних тому, що зовнішні стимули лише запускають їх, напрямок руху залежить від внутрішніх факторів.
Фотонастичні рухи виникають внаслідок зміни інтенсивності освітлення. Квітки кульбаби на ніч закриваються, а вранці розкриваються. Протилежне явище спостерігається у рослин, у яких на світлі квітки закриваються, а при зменшенні інтенсивності освітлення розкриваються. Такі рухи відрізняються від тропізмів, що пов’язані з ростом.
Листкові пластинки рослин, здатних до фотонастії, вдень розміщуються горизонтально, тоді як ввечері змінюють своє положення шляхом викривлення черешків чи листкових пластинок.
Термонастії виникають внаслідок зміни температури середовища. Такий рух можна спостерігати тоді, коли квітки тюльпана, шафрана перенести з холоду в умови 20-25’С. У таких умовах квітки названих рослин розкриваються.
У багатьох рослин, особливо бобових (конюшина, мімоза), в листках та листочках є особливі структури — листкові подушечки. Це особливе здуття при основі черешка або листочка, де знаходяться великі паренхімні клітини. Швидка зміна тургорного тиску в таких клітинах приводить до того, що листкова подушечка функціонує як шарнір, за допомогою якого і відбуваються рухи.
Гіпонастичні рухи, які спостерігаються у відповідь на Дотик, належать до найбільш специфічних рухів, властивих рослинам. Всім відома чутлива рослина мімоза соромлива (Mimosapudica), яка досить чутлива на дотик. У цього виду є і звичайні сонні рухи, але він здатний різко реагувати на "шокові» впливи — сейсмонастіі. Подразнення кінчиків листочків зумовлює згортання їх уже через кілька секунд. Більша сила впливу чи тривалість дії подразника може спричинювати не лише попарне згортання листочків, а і явище поникання всього черешка (рис. 207).
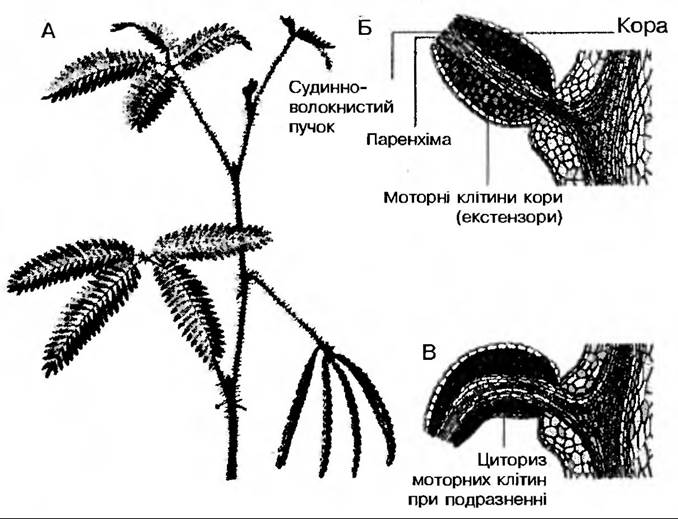
Рис. 207. Настичні рухи Mimosa pudica
Вважають, що механізм сейсмонастичних рухів зумовлюється зміною тургору верхньої та нижньої половини зчленування, яка підтримує листок у піднятому стані. Це пов’язано із зміною транспортування іонів в локомоторних клітинах. Моторні клітини взаємодіють з іншими клітинами кори та клітинами коленхіми подушечки. Аналогічно продиховим клітинам зміна іонних потоків пов’язана з роботою протонних помп, які виносять Н+ з моторних клітин одночасно з транспортуванням К+, СІ- та інших іонів. В процесі руху взаємодіють протонні помпи моторних клітин верхніх та нижніх частин подушечки, які функціонують у протилежних фазах. Як вважають, подразнення передає фітогормон, який переноситься по ксилемі. Передача сигналу пов’язана також з електричними змінами. Доведено, що подразнення спричинює появу в листку потенціалів дії з амплітудою 50-100 мВ, які поширюються по черешку із швидкістю 0,5-4 см/с. Через 0,05-1 с після появи потенціалу дії в подушечці електричний опір нижніх моторних клітин знижується порівняно з верхніми за рахунок виходу іонів калію та хлору із цих клітин. Вихід іонів з вакуолі супроводжується виходом води в міжклітинний простір, що призводить до втрати тургору клітинами. Допускають також участь білкових скоротливих систем моторних клітин у швидких сейсмонастичних рухах деяких рослин.
У комахоїдної рослини венерина мухоловка (Dionaea muscipula) листки, які посаджені на крилаті черешки, швидко складаються, коли комаха зачіпає чутливі волоски на верхній стороні листка. Вважалось, що в цих рухах теж бере участь тургор. Але в останні роки було виявлено, що швидке закривання листків венериної мухоловки пов’язане з незворотним розтягуванням клітин, яке розпочинається з підкислення клітинних стінок до pH 4,5 одиниці і нижче. У міру закривання клітини епідерми нижньої центральної частини кожного листка частково розтягуються, тоді як розміри клітин верхньої епідерми залишаються майже без змін.
Навпаки, при відкриванні листків клітини верхньої епідерми розтягуються, а нижньої залишаються без змін. Контроль рівня АТФ в період закривання листка показав, що третина наявного АТФ витрачається за цей короткий проміжок (1-3 с) часу. Припускають, що АТФ витрачається для швидкого транспортування водню. Є й інші форми сейсмонастичних реакцій. Всі рослини, які реагують на дотик, струс, реагують і на електричний струм. Рухи листочків мімози можна спричинити і пораненням певного листочка, чи всієї рослини — травмонастії. Два окремі механізми (електричний і хімічний) завжди включаються при поширенні стимулу в таких рослинах.
Настичні рухи продихового апарата. Відкривання та закривання продихового апарата здійснюється завдяки настичним тургорним рухам замикаючих клітин. Для продихів характерні фото-, гігро-, термо- та хемонастії. Рухи клітин продихового апарата — єдина форма рухів, при яких зміна тургору виникає через зміну осмотичного потенціалу. Це активні процеси, які відбуваються із затратами АТФ для міжклітинного транспортування іонів, особливо К+.
Різноманітні для кожного виду рослин продихи влаштовані таким чином, що при високому тургорі вони відкриваються і закриваються при низькому. Морфологічною основою рухів продихового апарата є структура замикаючих бобоподібних клітин. Зовнішні стінки цих клітин еластичні, а внутрішні потовщені, нееластичні. Рух у певному напрямку підтримується завдяки особливій мікроструктурі та розміщенню мікрофібрил внутрішньої стінки. Ступінь відкриття продихів є функцією різниці тургорного тиску замикаючих клітин та сусідніх клітин епідермісу, які чинять опір їхньому розтягуванню. Спектр дії фотонастичних рухів продихів подібний до спектра дії фотосинтезу, хоча висока активність синього світла дозволяє припустити, що в цьому процесі, крім фотосинтетичних, беруть участь і інші пігменти.
Не виключено, що індуковані зміною день — ніч рухи продихів здійснюються завдяки перетворенням крохмалю в цукри, а короткочасні зміни ширини продихового отвору під впливом різних факторів — результат включення в дію іонних насосів. Замикаючі клітини також реагують на вміст СО2 в міжклітинному просторі (при концентрації приблизно 0,05% СО2 вони закриваються, нижчій — відкриваються). Специфічний ефект синього світла, циркадний ритм у відкриванні продихів, ймовірно, взаємозв’язані із зміною концентрації СО2.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.