БІОФІЗИКА РОСЛИН - Ю. І. Посудін - 2004
IІ. ПРОЦЕСИ ПЕРЕНОСУ В СИСТЕМІ РОСЛИНА-ҐРУНТ-ПОВІТРЯ
6. ПЕРЕНОС МАСИ
6.8. ПОТОКИ ВОДИ ТА РОЗЧИНЕНИХ РЕЧОВИН У РОСЛИНАХ
6.8.1. Потік води в клітині
Різниця водних потенціалів ΔΨв пo обидві сторони мембрани або мембран, що обмежують об'єм потоку води Jk, є рушійною силою, яка забезпечує цей потік:
![]()
де Jв - об’єм потоку води, що протікає через одиницю поверхні за одиницю часу (м3/м2·с = м·с-1), Lв - коефіцієнт провідності води (м·с-1·МПа-1), Ψев та Ψів — водні потенціали зовнішнього та внутрішнього розчинів відповідно.
Якщо розглядати водний потік через кілька (j) клітин, послідовно з’єднаних одна з одною, то процес переносу води описується таким рівнянням:
![]()
Розрахуємо різницю водних потенціалів, необхідну для забезпечення потоку води, що спостерігається у прісноводних водоростях Nitella та Chara, клітини міжвузля яких характеризуються довжиною l = 10 см та діаметром r = 1 мм. Коефіцієнт провідності води для таких клітин дорівнює Lв = 10-6 м·с-1·МПа-1. Вода проходить із зовнішнього середовища через бокову поверхню клітин, площа якої становить ![]() Об’єм циліндра, який утворює Клітина, дорівнює
Об’єм циліндра, який утворює Клітина, дорівнює ![]() Припускаючи, що швидкість зростання клітини (та відповідне збільшення вмісту води) становить 1 % від об’єму клітини за добу, оцінимо загальний об’єм потоку води протягом дня
Припускаючи, що швидкість зростання клітини (та відповідне збільшення вмісту води) становить 1 % від об’єму клітини за добу, оцінимо загальний об’єм потоку води протягом дня ![]() Звідси швидкість надходження води через одиницю площі бокової поверхні клітини за одиницю часу дорівнює.
Звідси швидкість надходження води через одиницю площі бокової поверхні клітини за одиницю часу дорівнює.
![]()
Отже, знаходимо різницю водних потенціалів ΔΨв з рівняння (6.22).
![]()
Розглянуті водорості мешкають у ставовій воді, яка є розведеним водним розчином, водний потенціал якого дорівнює -0,007 МПа. Отже, водний потенціал у клітині має бути на 3·10-5 МПа меншим, ніж водний потенціал зовнішнього середовища, з тим щоб забезпечити приплив води у клітину.
6.8.2. Потік розчинених речовин у клітині
Розглянемо випадок спряження двох потоків через мембрану - води та розчиненої речовини. Мембрана проникна і для води, і для розчиненої речовини. Рушійною силою обох потоків є градієнти хімічного потенціалу. Використовуючи рівняння (6.22), представимо кожний потік як лінійну комбінацію рушійних сил:
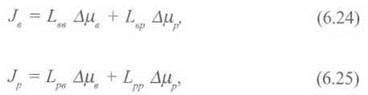
де індекси в та р стосуються води і розчину відповідно.
З рівнянь видно, що кожний потік залежить від обох хімічних потенціалів. Оскільки згідно з принципом симетрії феноменологічних коефіцієнтів Онзагера Lвр = Lрв, для опису залежності двох потоків від двох рушійних сил потрібно знати лише три феноменологічні коефіцієнти.
Різницю хімічних потенціалів можна подати через осмотичний та гідростатичний тиски. Використовуючи рівняння (6.13), знайдемо різницю хімічних потенціалів:
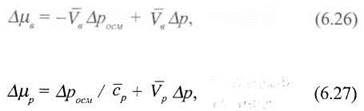
де ![]() - середня концентрація розчиненої речовини.
- середня концентрація розчиненої речовини.
Таким чином, зміни хімічних потенціалів Δμв, та Δμр, які є рушійними силами процесу, подані як функції тих самих різниць тисків Δросм та Δр.
6.8.3. Потік води у системі ґрунт-рослина-атмосфера
Вода є основним середовищем, в якому відбуваються біохімічні та біофізичні процеси, що забезпечують метаболізм рослини: розчиняє цукри та мінеральні речовини, які транспортуються через рослину, виступає як реагент у процесі фотосинтезу, підтримує необхідний тургорний тиск у клітинах, забезпечує потрібну температуру в рослині за рахунок випаровування, зменшує густину оточуючого повітря і сприяє тим самим циркуляції повітря у рослинних покривах. Вода рухається безперервним потоком з ґрунту через кореневу систему і стебло рослини до листя і далі, через продихи, в оточуючу атмосферу. Це безперервний, але неоднорідний потік, оскільки протягом свого руху вода переміщується у ґрунті до коріння, поглинається кореневою системою, транспортується по судинах ксилеми, дифундує через міжклітинні проміжки та продихи і, врешті-решт, у вигляді водяної пари залишає листя і виходить в атмосферу. Рушійною силою, що забезпечує рух води, є: у ґрунті - градієнт тиску, в кореневій системі - Градієнт водного потенціалу, транспортування води на велику відстань по ксилемі - градієнт тиску, під час транспірації - градієнт концентрації водяної пари (рис. 6.14).
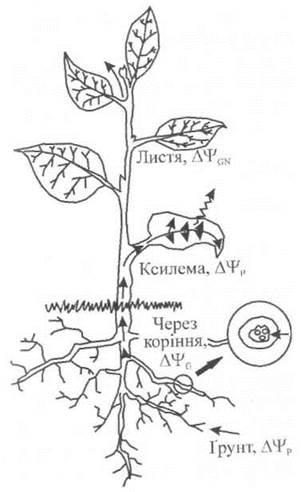
Рис. 6.14. Основні рушійні сили, що забезпечують Рух води по системі "ґрунт-рослина- атмосфера": у ґрунті - градієнт тиску, в кореневій системі - градієнт водного потенціалу, транспортування води на велику відстань по ксилемі - градієнт тиску, під час транспірації - градієнт концентрації водяної пари.
Кількість води, що виходить з рослини завдяки транспірації, дорівнює тій кількості води, яка поглинається кореневою системою. Отже, водний потік у рослині можна вважати постійним. За цими умовами можна застосовувати для водного потоку аналогії з електричного ланцюга - водний потік розглядати як електричний струм, який дорівнює відношенню різниці водних потенціалів ΔΨ до опору г переносу води:
![]()
Застосовуючи останнє рівняння до системи «ґрунт-рослина-атмосфера», отримаємо:
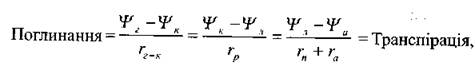
де Ψг - водний потенціал ґрунту, Ψк - водний потенціал коріння, Ψл — водний потенціал листка, Ψа - водний потенціал атмосфери, rг-к — опір проходженню води з ґрунту у кореневу систему, rp - внутрішній опір рослини, rn - опір продихів, ra - опір атмосфери (граничного шару).
Перші два відношення відповідають потокам рідкої фази, тоді як останній - потоку газової фази. Типові різниці водних потенціалів, що спостерігалися для різних ділянок системи ґрунт—рослина—атмосфера, такі [Cruiziat, 1987]:
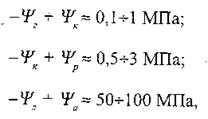
де Ψг, Ψк, Ψp, Ψл, Ψа - водні потенціали ґрунту, коріння, рослини в цілому, листя та водяної пари в атмосфері відповідно.
Зменшення водного потенціалу протягом транспортування води в системі ґрунт-рослина-атмосфера наведено на рис. 6.15. Водний потенціал вологого ґрунту, кореневої системи та стебла близький до нуля, що відповідає його максимальному значенню (крива 1). Листя характеризується меншим водним потенціалом, якого все ж таки достатньо для запобігання в'яненню; продихи в такій ситуації відкриті. Зменшення водного потенціалу ґрунту (крива 2) викликає відповідне зменшення водного потоку, водного потенціалу листків, завдяки чому продихи закриваються.
Водний потенціал вологого ґрунту складається з осмотичного потенціалу та потенціалу тиску. Осмотичний потенціал ґрунтової води достатньо малий (-0,02 МПа), через що ним можна знехтувати, хоча в солонуватих ґрунтах осмотичний потенціал збільшується до -0,2 МПа. Гідростатичний тиск вологого ґрунту близький до нуля. Підсихання ґрунту викликає видалення води з проміжків між ґрунтовими частинками та утворення угнутих поверхонь - менісків. Напружений стан поверхневого шару рідини називається поверхневим натягом, а сили, які зумовлюють скорочення поверхневої плівки рідини, називають силами поверхневого натягу. Вода характеризується надзвичайно високим поверхневим натягом: при 20 °С поверхневий натяг на межі поділу повітря-вода дорівнює 72,8·10-3 Н·м-1. Якщо поверхня рідини викривлена, то вона здійснює на рідину додатковий тиск, обумовлений силами поверхневого натягу. Цей додатковий тиск для сферичної викривленої поверхні визначається за формулою Лапласа:
![]()
де σ- коефіцієнт поверхневого натягу, R - радіус кривизни поверхні рідини. Додатковий тиск береться зі знаком якщо поверхня опукла, зі знаком якщо поверхня угнута. Отже, на межі поділу вода-повітря виникає від’ємний додатковий тиск Ψp = -2σ/R, де R - радіус меніска.
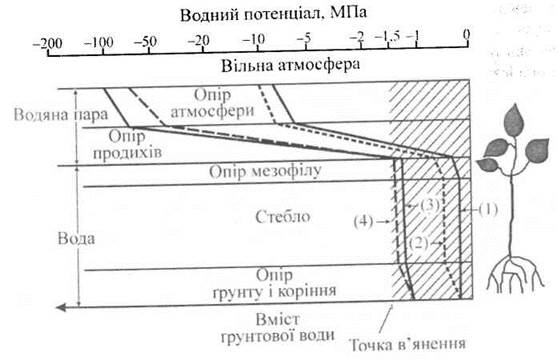
Рис. 6.15. Зменшення водного потенціалу протягом транспортування води в системі "ґрунт-рослина-атмосфера".
У попередніх розділах зазначалося, що основними механізмами переносу води є молекулярна дифузія та об'ємний потік. На великих відстанях переносом води керує об'ємний потік, який викликається градієнтом тиску. Завдяки поглинанню води рослиною ґрунтова вода вичерпується, що призводить до зменшення потенціалу тиску Ψp води поблизу поверхні кореневої системи. Це призводить до утворення градієнта тиску, вздовж якого вода з інших ділянок ґрунту рухається через ґрунтові пори. Швидкість водного потоку залежить не лише від величини градієнта тиску, але й від гідравлічної провідності ґрунту, яка залежить від типу ґрунту (пісок, наприклад, має високу гідравлічну провідність, чого не можна сказати про глину). В занадто сухих ґрунтах водний потенціал Ψ може спадати до граничної величини, яка отримала назву точки в’янення. В такій ситуації рослина неспроможна оновлювати тургорний тиск і в'яне.
6.8.4. Коренева система
Поглинання рослиною води та мінеральних поживних речовин із ґрунту здійснюється за допомогою кореневої системи. Надійний контакт коріння з ґрунтом забезпечується завдяки великій кількості коренів та кореневих волосків у рослини. Наприклад, можна навести параметри кореневої системи чотиримісячної рослини жита [Полевой, 1989]: середня кількість коренів - 1,38 · 107; загальна площа кореневої системи - 232 м2; кількість кореневих волосків - 1,4 · 1010; сумарна площа коренів та кореневих волосків - 631 м2. Причому сумарна площа поверхні кореневих волосків може досягати 60 % загальної площі кореневої системи.
Протягом свого руху через кореневу систему вода, поглинута кореневим волоском, перетинає епідерміс і транспортується або симпластним шляхом - через цитоплазму по плазмодесмах, які являють собою найдрібніші цитоплазматичні тяжі, що сполучають протопласти сусідніх клітин, або апопластним шляхом - через клітинні стінки (рис. 6.16).
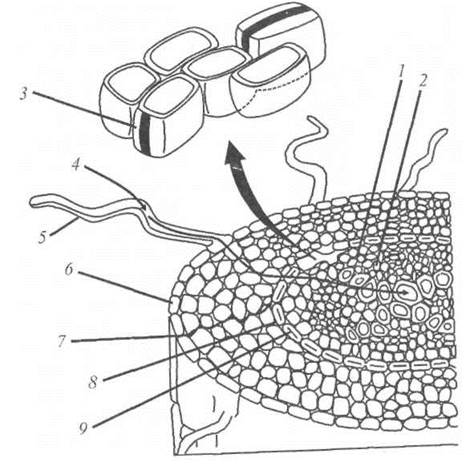
Рис. 6.16. Шляхи транспортування води через кореневу систему симпластним або апопластним шляхом: 1 - флоема, 2 - ксилема 3 - поясок Каспарі 4 - клітинний шлях, 5 - кореневий волосок, 6 - епідерміс, 7 - апопластний шлях, 8 - ендодерма, 9 - поясок Каспарі.
Є гіпотеза [Fensom, 1975; Spanner, 1975], яка передбачає можливість транспортування речовин по кореневій системі завдяки електроосмосу - руху рідини вздовж стінок капілярів, системи капілярів або пористих пробок під впливом зовнішніх електричних полів. У макроскопічній однорідній системі вода нечутлива до цих полів завдяки своїй електронейтральності. На межі поділу стінки капіляра з рідиною утворюється подвійний шар електричних зарядів - нерухомих, які поглинуті стінкою, та рухомих, що знаходяться в рідині. Рухомі заряди сприяють виникненню на ситоподібних пластинках електричного поля. Швидкість руху електричних зарядів визначається за виразом [Ksenzhek, Volkov, 1998]:
![]()
де ε - діелектрична проникність середовища (для води 80), εд — діелектрична проникність вакууму (8,85 · 10-12 Кл·В-1·м-1), С - електрокінетичний потенціал, який являє собою різницю потенціалів між нерухомими та рухомими зарядами і становить від кількох одиниць до десятків мілівольт, η - в'язкість, Е - напруженість електричного поля (В·м-2).
Швидкість електроосмотичного потоку невелика - для типових значень ξ = 20 мВ та Е = 104 В·м-1 швидкість V = 1,4 · 102 см·с-1. Оскільки ця швидкість не залежить від радіуса капіляра, можна припустити, що цей механізм відіграє певну роль у капілярах малого діаметра. Є дані [Dainty, 1963], які свідчать про можливість виникнення градієнта тиску в кореневій системі близько 1 МПа при різниці потенціалів 10 мВ.
6.8.5. Ксилема
Систему, що забезпечує постачання води та мінеральних речовин з ґрунту у верхні частини рослини, називають ксилемою. Ця тканина складається з провідних елементів - трахеїд, волокон та елементів судин, що утворюють довгі ряди вздовж розташованих клітин. Діаметри провідних елементів ксилеми варіюють у діапазоні 10-500 мкм для різних видів рослин, а довжина може становити від декількох сот мікрометрів до двох і більше метрів. Через те, що ці провідні елементи позбавлені протопластів і клітинних оболонок, опір проходженню через них води та розчинів дуже малий. Спрощений вигляд провідного елемента ксилеми наведено на рис. 6.17, а.
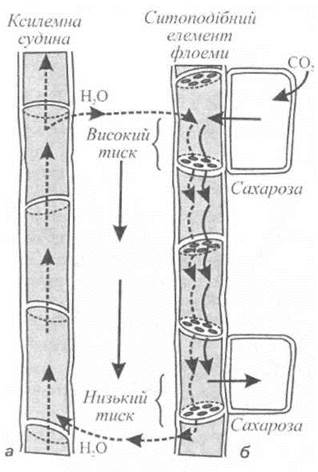
Рис. 6.17. Спрощений вигляд провідних елементів рослини: а - ксилеми; б - флоеми.
6.8.6. Механізми ксилемного транспорту
Якщо помістити вузьку трубку (капіляр) одним кінцем у рідину, то внаслідок змочування або незмочування рідиною стінок капіляра кривизна поверхні рідини (меніска) стає суттєвою, через що над поверхнею утворюється значний додатковий тиск. Притягання між молекулами рідини називають когезією, тоді як притягання між рідиною і твердим тілом (стінками капіляра) - адгезією. Коли взаємодія рідини зі стінкою виявляється значною, відбувається змочування стінки рідиною; коли ж міжмолекулярні когезійні сили в рідині значно перевищують адгезію між рідиною та стінкою, має місце незмочування стінки рідиною. Розглянемо капіляр, занурений у змочуючу рідину. Сили притягання, які виникають між молекулами рідини і капіляра, примушують рідину підійматися по стінці капіляра, що призводить до викривлення поверхні рідини і утворення негативного тиску. Внаслідок цього рідина підіймається по капіляру, доки гідростатичний тиск не зрівноважить додатковий тиск. Умову рівноваги можна описати за виразом:
![]()
де ρ - густина рідини, R - радіус кривизни меніска, g - прискорення вільного падіння, h - висота, на яку підіймається рідина. Звідси можна визначити висоту піднімання рідини:
![]()
де r = Rcosθ - радіус капіляра, θ - крайовий кут (рис. 6.18).
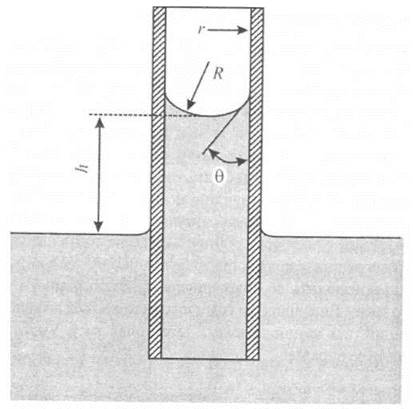
Рис. 6.18. Величини, необхідні для розгляду капілярного підіймання води.
Розглянемо на конкретному прикладі судину ксилеми радіусом 20 мкм: згідно з формулою (6.32) вода в судині підійметься на висоту:
![]()
Отже, додатковий тиск у судинах ксилеми не здатний забезпечити піднімання рідини в рослинах висотою більше одного метра. На великих відстанях переносом води керує об'ємний потік, який забезпечується градієнтом тиску.
Кількісно цей об'ємний потік води J для циліндричних трубок, таких як клітини ксилеми радіусом г, описується рівнянням Пуазейля:
![]()
де η - в’язкість рідини, ![]() — градієнт тиску. З рівняння видно, що об'ємний потік суттєво залежить від радіуса трубки: якщо збільшити радіус удвічі, швидкість об'ємного потоку води збільшиться у 24 = 16 разів. Таким чином, керований градієнтом тиску об'ємний потік води є домінуючим механізмом руху води на великі відстані.
— градієнт тиску. З рівняння видно, що об'ємний потік суттєво залежить від радіуса трубки: якщо збільшити радіус удвічі, швидкість об'ємного потоку води збільшиться у 24 = 16 разів. Таким чином, керований градієнтом тиску об'ємний потік води є домінуючим механізмом руху води на великі відстані.
Застосуємо рівняння Пуазейля для оцінки об'ємної швидкості протікання води через одиницю площі (S = nr2) судини ксилеми, який у цьому разі визначається за виразом:
![]()
Якщо середній потік води в ксилемі становить 10-3 м·с-1, коефіцієнт в’язкості води η = 10-3 Па·с, а радіус ксилеми r = 20 мкм = 2·105 м, то градієнт тиску, що задовольняє рівняння (6.33), становитиме = -2·104 Н·м3 = -2·104 Па·м-1.
Зміна гравітаційного потенціалу на кожний метр висоти становить (як було встановлено в попередніх розділах) ρgh/h = 104 Па = 10-2 МПа. Тобто, градієнт тиску спроможний перевищити силу тяжіння і підтримати рух води в ксилемі.
6.8.7. Флоема
Під час фотосинтезу відбувається перетворення світлової енергії на хімічну, яке супроводжується утворенням вуглеводів з вуглекислого газу. Хоча невелика кількість сполук вуглецю або фотоасимілятів іде на забезпечення процесів росту та метаболізму листя, основна кількість передається нефотосинтетичним органам та тканинам. Процес транспортування фотоасимілятів на довгі відстані називається транслокацією. Розподіл більшості органічних речовин (у першу чергу - фотоасимілятів) по рослині здійснюється завдяки флоемі - другій провідній системі рослини. Флоема являє собою тканину, що складається з кількох типів клітин. Але на відміну від ксилеми провідні елементи флоеми містять протоплазму (рис. 6.17, б). Ці провідні елементи називають ситоподібними клітинами або ситоподібними трубками. Вони з'єднані в єдиний ланцюг, який забезпечує транспортування органічних речовин. Розчинені речовини переміщуються по флоемі - продукти фотосинтезу рухаються від листя до кореневої системи, тоді як цукри у протилежному напрямку, від кореневої системи до верхівки рослини.
6.8.8. Механізми транслокації по флоемі
Будь-яка теорія, яка б могла пояснити механізми транслокації фотоасимілятів та органічних речовин по флоемі, повинна враховувати та пояснювати структуру ситоподібних елементів флоеми, високу (50—250 см·г-1) швидкість транслокації на великі відстані, одночасне транспортування речовин у протилежних напрямках, початковий процес перенесення асимілятів з мезофільних клітин листка до ситоподібних елементів флоеми (завантаження флоеми), транспортування асимілятів по ситоподібних елементах та остаточне перенесення асимілятів із ситоподібних елементів до клітин запасаючих органів (розвантаження флоеми).
Найбільш вірогідною вважається гіпотеза потоку під тиском[Miinch, 1930], яка базується на переносі маси розчиненої речовини від джерела до запасаючих органів за рахунок градієнта гідростатичного (тургорного) тиску в ситоподібних елементах (див. рис. 6.17). Завантаження флоеми відбувається завдяки транспортуванню цукрів від мезофілу через тонкі пучки, серединну жилку та листовий черешок до ситоподібних елементів флоеми. Збільшення концентрації розчинених речовин у цих елементах призводить до зменшенню водного потенціалу та відповідного осмотичного поглинання води із сусідньої ксилеми. Гідростатичний (тургорний) тиск на цьому кінці флоеми збільшується. У той же час на протилежному кінці флоеми цукри покидають флоему, а вода повертається у ксилему. Тиск зменшується, а в цілому вздовж флоеми утворюється градієнт тиску, який обумовлює потік рідини від листя до коріння. Цей градієнт описується законом Пуазейля. На користь цієї гіпотези свідчать результати експериментального вимірювання гідростатичного тиску в ситоподібних елементах, який становить 2·106 Па[Нобел, 1973].
Ще одна гіпотеза [Fensom, 1975; Spanner, 1975] передбачає участь електроосмосу у транспортуванні речовин по флоемі (див. розділ 6.8.4 "Коренева система"). Вважається, що рушійною силою переміщення цукрів з однієї ситоподібної трубки в іншу через пори є транспорт іонів ІC, який супроводжується виникненням електричних полів на кожній ситоподібній пластинці. Існують ще гіпотези, що пояснюють механізми транспортування асимілятів по ситоподібних елементах - рух протоплазми, перистальтика (скорочення) клітинних стінок, мікроелектрокінез, поверхнево-активний рух та інші, але вони недостатньо переконливі [Люттге, Хігінботам, 1984].
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.