БІОФІЗИКА РОСЛИН - Ю. І. Посудін - 2004
IІ. ПРОЦЕСИ ПЕРЕНОСУ В СИСТЕМІ РОСЛИНА-ҐРУНТ-ПОВІТРЯ
6. ПЕРЕНОС МАСИ
6.7. ВИМІРЮВАННЯ ВОДНОГО ПОТЕНЦІАЛУ ТА НОГО КОМПОНЕНТІВ
6.7.1. Метод реєстрації зміни ваги Тканини
Водний потенціал рослинної тканини можна оцінити шляхом досягнення рівноваги між осмотичним потенціалом розчину та водним потенціалом тканини. Для цього заздалегідь зважений зразок тканини занурюють у ємність з розчином, осмотичний потенціал якого відомий. Якщо осмотичний потенціал буде більш від'ємним, ніж водний потенціал тканини, Вода залишить тканину, яка позбавиться внаслідок цього певної ваги; якщо ж осмотичний потенціал розчину буде менш від'ємним, ніж водний потенціал тканини, вода буде надходити у тканину і вага останньої збільшиться. Розчин, який не спричиняє втрату чи надбавку ваги, еквівалентний водному потенціалу тканини.
На практиці процедура вимірювань (рис. 6.2) складається з підготовки зразків, однакових за розміром та масою, які занурюють у розчин відомої моляльності (згадаємо, що кількість молів на 1 л розчину називають молярністю, а на 1 кг розчину - моляльністю). Для того, щоб запобігти зміні концентрації водного розчину протягом вимірювання, використовують такі речовини, що не поглинаються тканиною (сорбітол, манітол, поліетилен-гліколь). Через тривалий проміжок часу настає рівновага між тканиною та розчином, після чого тканину позбавляють залишків розчину і знову зважують. Будують графік залежності зміни ваги від концентрації розчину (рис. 6.3). Використовуючи рівняння Вант-Гоффа, визначають осмотичний потенціал і, отже, водний потенціал.

Рис. 6.2. Метод реєстрації зміни ваги тканини (пояснення в тексті).
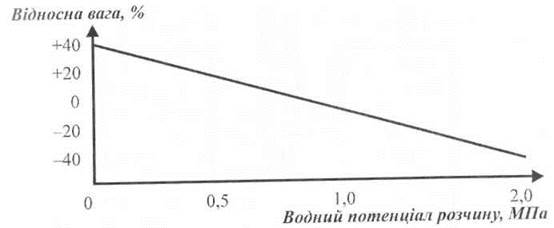
Рис. 6.3. Графік залежності зміни ваги від водного потенціалу розчину.
6.7.2. Термопарна психрометрія
Маленький зразок рослинної тканини (листка) розміщують у камері, яка обладнана температурним сенсором - термопарою. Внаслідок випаровування води з поверхні листка Температура останнього зменшується. Оскільки вода залишає рослинну тканину, водний потенціал останньої зменшується, доки не настане стан рівноваги. Коли водна пара і рослинна тканина знаходяться в рівновазі, вони мають однакові значення водного потенціалу. Оцінити водний потенціал можна шляхом вимірювання тиску водяної пари в камері, який пов'язаний з температурою. Процес вимірювання температури базується на використанні двох термопар, що утворюють електричний ланцюг. Спай однієї термопари знаходиться в камері, тоді як спай іншої пов'язаний із джерелом опорної температури. Якщо спаї двох термопар мають різну температуру, в електричному ланцюзі виникає електрорушійна сила і через ланцюг починає текти електричний струм. Це термоелектричне явище отримало назву ефекта Зеєбека.
Метод термопарної психрометрії передбачає використання термопари, обладнаної срібним кільцем, на яке нанесена водна крапля (рис. 6.4). Спочатку відбувається процес випаровування води з поверхні листка та водної краплі. Оскільки камера мала за розмірами, внутрішня атмосфера швидко досягає стану насичення. Якщо рослинна тканина та водна крапля мають однакові значення водного потенціалу, сумарний потік води з краплі припиняється, і температура краплі стає рівною оточуючій температурі. Але якщо водний потенціал рослинної тканини більш від'ємний, ніж потенціал водної краплі, вода дифундує з атмосфери у тканину. Процес випаровування з поверхні краплі, направлений на компенсацію тієї водяної пари, що поглинута тканиною, супроводжується охолодженням термопари, на якій знаходиться крапля. Швидкість переносу маси за рахунок дифузії пропорційна різниці значень водного потенціалу краплі та тканини. Отже, вимірювання швидкості охолодження термопари дає можливість оцінити тиск водної атмосфери в камері та водний потенціал тканини. Недоліком способу є високе значення дифузійного опору переносу води з боку рослинної тканини, що викликає похибки вимірювань.
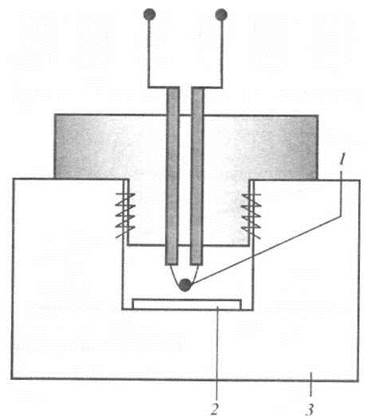
Рис. 6.4. Суть методу термопарної психрометрії: 1 - термопара; 2 - зразок; 3 — термостат.
Модифікація методу термопарної психрометрії (ізопієстична техніка) відрізняється тим, що замість води на спай термопари наносять краплину розчину відомої концентрації і, отже, відомого водного потенціалу [Boyer and Rnipling, 1965]. Якщо водний потенціал розчину перевищує водний потенціал рослинної тканини, вода буде переноситися з розчину у тканину, що викликає охолодження термопари. У випадку, коли водний потенціал розчину менший за водний потенціал рослинної тканини, відбувається зворотний процес переносу води з тканини у розчин. Цей процес супроводжується конденсацією води на термопарі з подальшим збільшенням її температури. Зрозуміло, що коли водні потенціали розчину та тканини однакові, тиски водяної пари поблизу термопари та тканини зрівноважуються і процес охолодження термопари внаслідок випаровування припиняється. Температура термопари в цьому разі досягає температури навколишнього середовища. На практиці використовують два розчини з відомою концентрацією і вимірюють сигнали термопари для кожного розчину. Оскільки залежність між значеннями водного потенціалу та вихідного сигналу термопари лінійна, можна провести пряму лінію між отриманими двома точками. Екстраполяція цієї прямої в область, де вихідний сигнал термопари дорівнює нулю, дає можливість визначити ізопієстичну точку, яка свідчить про припинення руху водяної пари між розчином та термопарою і відповідає водному потенціалу тканини. Цей метод характеризується високою чутливістю та дозволяє вимірювати водні потенціали з розділенням близько 0,01 МПа. До недоліків методу можна віднести необхідність дуже ретельного підтримання температури: зміна температури на 0,01 °С супроводжується відповідною зміною водного потенціалу на 0,1 МПа. Саме тому метод використовують лише у лабораторних умовах, а не в польових.
6.7.3. Камера тиску
Цей простий, але швидкодіючий метод базується на вимірюванні від'ємного гідростатичного тиску в ксилемі [Scolander et al., 1965]. Вважається, що водний потенціал ксилеми приблизно дорівнює середньому водному потенціалу всієї рослини. Це припущення вірне, оскільки осмотичним потенціалом розчину в ксилемі можна знехтувати, через що основним компонентом водного потенціалу в ксилемі є від'ємний гідростатичний тиск; крім того, ксилема знаходиться у тісному контакті з більшістю клітин рослини. Для реалізації методу листок чи паросток відокремлюють від рослини і фіксують у камері тиску (рис. 6.5). Причому, вода покидає місце зрізу завдяки ксилемному тиску. Для того, щоб повернути воду на це місце, в камері збільшують внутрішній тиск за допомогою джерела стиснутого газу доти, поки вода знову не з'явиться на місці зрізу. Цей тиск балансування дорівнює за величиною (але з протилежним знаком) негативному гідростатичному тиску в ксилемі, який був у зразку перед відокремлюванням.
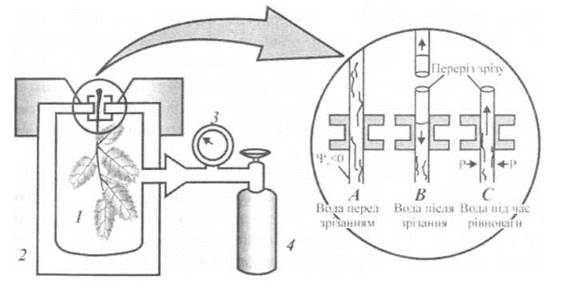
Рис. 6.5. Фіксація листка чи паростка в камері тиску: 1 - зразок, 2- камера тиску, 3 - манометр, 4 - балон з газом.
6.7.4. Метод ядерного магнітного резонансу
Більшість методів вимірювання водного потенціалу вимагає руйнування зразків. Внаслідок цього дуже важко слідкувати за динамічними процесами переносу води в тканині протягом часу. Ці недоліки відсутні у техніці ядерного магнітного резонансу, в основі якої лежить селективне поглинання електромагнітної енергії речовиною, обумовлене квантовими переходами атомних ядер між енергетичними станами з різними орієнтаціями спіну ядра.
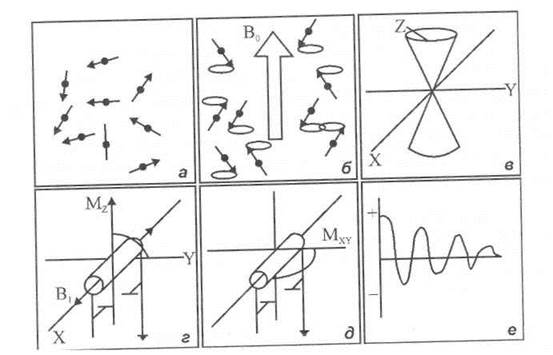
Рис. 6.6. Поведінка дипольних моментів ядер у зовнішньому магнітному полі: а - дипольні моменти ядер; б - прецесія диполів навколо напрямку зовнішнього статичного магнітного поля; в - прецесія двох спінів у протилежних напрямках; г - поява сумарної намагніченості завдяки різниці орієнтацій двох груп диполів; д - обертання вектора Мz на 90° при забезпеченні п/2-імпульсу тривалості радіочастотного магнітного поля; е - форма затухаючої косинусоїдної хвилі сигналу, що реєструється.
Відомо, що ядра всіх елементів мають електричний заряд, який є додатним і дорівнює за абсолютною величиною сумі зарядів атомних електронів. Завдяки власному моменту імпульсу (спіну) Ядро під час обертання діє як елементарний магніт. Отже, ядро характеризується магнітним моментом, величина якого залежить від природи ядра. Ядра з парним числом протонів і парним числом нейтронів не мають спіну і магнітного моменту, тоді як ядра з парним числом протонів і непарним числом нейтронів мають спін і дипольний магнітний моменте. Магнітний момент характеризується величиною та напрямком (рис. 6.6, а). Якщо зразок розмістити в інтенсивному однорідному магнітному полі В9, всі диполі починають прецесувати навколо напрямку магнітного поля (рис. 6.6, б) з ларморівською частотою, яка визначається так:
![]()
де γ - гіромагнітне відношення.
Причому одна група диполів виявляє сумарну орієнтацію у напрямку магнітного поля, тоді як інша - проти поля (рис. 6.6, в). Слід відзначити, що у стані рівноваги кількість диполів, що орієнтуються вздовж поля, перевищує кількість протилежно орієнтованих диполів. Це можна пояснити тим, що в основному енергетичному стані, коли магнітні диполі орієнтуються у напрямку магнітного поля, енергія ядра менша, ніж у збудженому, для якого притаманна орієнтація, протилежна магнітному полю.
Приклад. Магнітний момент ядра поводить себе як механічна система - гіроскоп. При обертанні гіроскопа навколо власної осі прикладання зовнішньої сили F у напрямку X викликає обертання осі гіроскопа навколо цієї осі, а не навколо осі у, як здавалося би з першого погляду. Внаслідок дії зовнішньої сили гіроскоп буде демонструвати прецесію у гравітаційному полі (рис. 6.7).
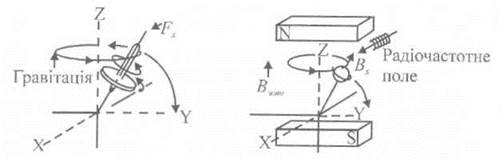
Рис. 6.7. Прецесія гіроскопа у гравітаційному полі.
Різниця енергій ΔЕ між рівнями пропорційна магнітній індукції:
![]()
це h - стала Планка.
Різниця населеностей основного та збудженого рівнів визначається за виразом:
![]()
де Nзб та N0 -населеності збудженого та основного рівнів відповідно, Т - температура.
Приклад. Визначити ларморівську частоту протона у магнітному полі, різницю енергій між збудженим та основним рівнями і відносну різницю населеностей, якщо магнітна індукція зовнішнього поля становить В0 = 2 Тл, гіромагнітне відношення протона γ = 42,6 МГц/Тл, а температура Т = 300 К.
Розв’язання. Ларморівську частоту можна визначити за виразом (6.14).
![]()
Різницю енергій між збудженим та основним рівнями визначимо за виразом (6.15).
![]()
Відносну різницю населеностей визначимо так:
![]()
За рахунок цієї різниці магнітних диполів утворюється сумарна намагніченість Мz, паралельна напрямку z постійного магнітного поля.
Якщо прикласти до зразка за допомогою котушок з електричним струмом зовнішнє радіочастотне магнітне поле В1, направлене перпендикулярно В0 (рис. 6.6, г), то воно буде взаємодіяти з магнітним моментом ядра μ. Ця взаємодія буде помітною, якщо частота ω радіочастотного поля буде близька до ларморівської частоти ω0. Якщо радіочастотне магнітне поле вмикають на кінцевий проміжок часу t, то вектор намагніченості Мz повернеться навколо В1 на кут θ = γB1t. Можна підібрати таку тривалість радіочастотного магнітного поля (π/2-імпульс), що вектор Мzобернеться на 90° (рис. 6.6, д). Після вимкнення радіочастотного магнітного поля В1 вектор намагніченості Мxy демонструє прецесію навколо осі z з частотою, що дорівнює різниці частот прецесій двох популяцій магнітних диполів — орієнтованих у напрямку поля та протилежно йому. Прецесія вектора намагніченості Мху фіксується відповідно до електричного струму, ЩО індукується в котушках індуктивності, вісь яких перпендикулярна напрямку зовнішнього магнітного поля В0. Сигнал, що реєструється, має форму затухаючої косинусоїдної хвилі (рис. 6.6, e).
Величини вектора намагніченості Мz та Мxy після вимкнення радіочастотного імпульсу змінюються з часом (рис. 6.8.): Мz зростає з часом за експоненціальним законом за виразом:
![]()
тоді як Мхy затухає за експоненціальним законом:
![]()
де T1 - час поздовжньої (спін-гратової) релаксації, Т2 - час поперечної (спін-спінової) релаксації, вимірювання яких є важливим діагностичним методом оцінки стану зразка.
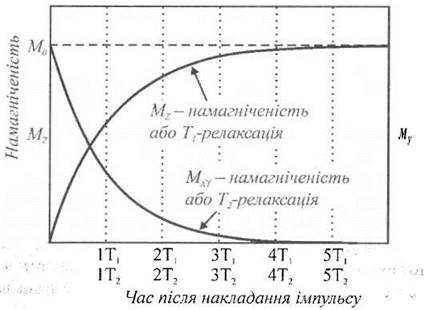
Рис. 6.8. Графік зміни величин векторів намагніченості Мz та Мxy з часом після вимкнення радіочастотного імпульсу.
Час поздовжньої релаксації T1 характеризує вплив ґратки - оточення ядра, яке складають решта молекули та молекули розчинника; час поперечної релаксації Т2 описує процес обміну енергією між сусідніми ядерними моментами без обміну енергією з ґраткою. Для вимірювання Т застосовують π/2-імпульс радіочастотного магнітного поля, тоді як дім вимірювання T2 доцільно прикладати π-імпульс, внаслідок чого вектор М, обернеться на 180° уздовж напрямку - z. Схему ЯМР-спектрометра наведено на рис. 6.9. Він складається з джерел статичного та радіочастотного магнітних полів, котушки індуктивності, вимірювальних приладів та зразка.
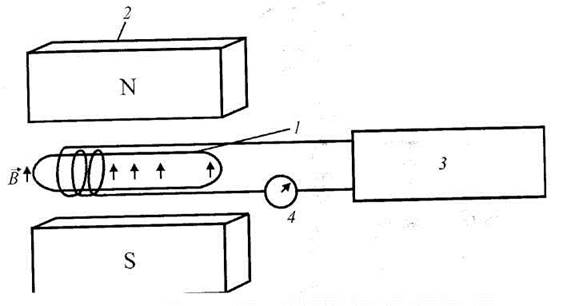
Рис. 6.9. Схема ЯМР-спектрометра: 1 - зразок, 2 - магніт, 3 - генератор, 4 - прилад.
Активність води в рослині обумовлюється тим, наскільки швидко молекули води беруть участь у хімічних реакціях або як легко молекули води дифундують у місця, де відбуваються ці реакції. Саме завдяки застосуванню техніки ЯМР стає можливим оцінити рухомість води в складних системах. Швидкі молекули води характеризуються довгим часом, протягом якого досягається стан рівноваги, або великими значеннями часів релаксації T1 або T2. У той же час, додавання у воду розчинника зменшує час релаксації (рис. 6.10). У біологічних молекулах час поперечної релаксації T2 залежить від можливих конформаційних змін молекули води, взаємодії води з мембранами, впливу процесів гідратації на білки. Отже, час поперечної релаксації протонів у клітинах може бути індикатором взаємодії клітинної води з мембранами або макромолекулами. Вимірювання цього параметра є ефективним неруйнівним методом оцінки фізіологічного стану води у клітинах.
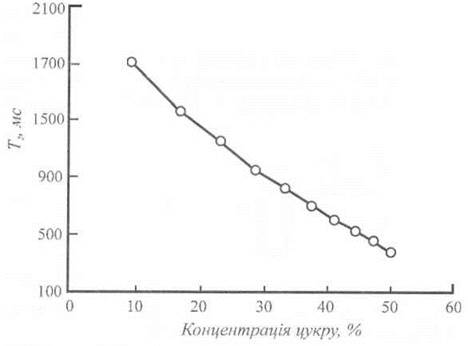
Рис. 6.10. Графік зменшення часу релаксації при додаванні у воду розчинника.
6.7.5. Вимірювання осмотичного потенціалу
Кріоскопічна осмометрія передбачає оцінку осмотичного потенціалу розчину при температурі замерзання. Однією з важливих властивостей розчинів є те, що температура їхнього замерзання залежить від концентрації розчиненої речовини. Наприклад, розчин, концентрація якого становить 1 моль·кг-1, замерзає при температурі -1,86 °С, тоді як у чистої води температура замерзання дорівнює 0 °С. Процедура вимірювань базується на мікроскопії зразків - розчинів концентрацією 1 нл (10-9 л), які розміщують в олії на предметному стільці, температура якого підтримується сталою (рис. 6.11). Спочатку спостерігають рослинний сік, який виходить із клітин; завдяки малим розмірам зразка стан терморівноваги досягається дуже швидко. Далі температуру зразка зменшують до -30 °С, при якій він замерзає. Після цього температуру повільно збільшують, фіксуючи через Мікроскоп процес плавлення зразка і вимірюючи температуру, при якій останні кристали льоду розплавляться (нагадаємо, що температура замерзання і температура плавлення однакові). Осмотичний потенціал оцінюють як Ψ = RTCс, де R - універсальна газова стала, Т - абсолютна температура, Сс - концентрація розчину. Ця техніка дозволяє працювати з краплинами, виділеними з поодиноких клітин.
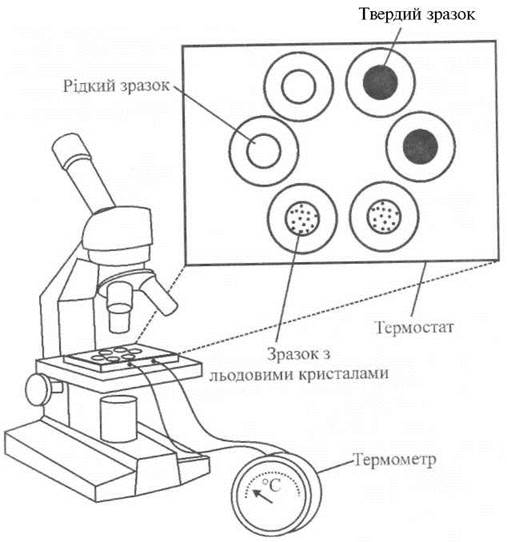
Рис. 6.11. Кріоскопічна осмометрія.
6.7.6. Вимірювання потенціалу тиску
Зонд тиску являє собою мікроманометр для вимірювання тургорного тиску в рослинних клітинах [Green and Stanton, 1967]. Скляний капіляр діаметром 2-7 мкм, з'єднаний з камерою тиску, заповнюють кремнієвою олією - нестисливою рідиною, яку легко відрізнити від води під мікроскопом. Зонд обладнаний плунжером, що забезпечує зміну тиску. Коли кінець капіляра занурюють у клітину (рис. 6.12), клітинний сік надходить у капіляр завдяки тургорному тиску. Після цього дослідник обертає ручку плунжера зонда так, щоб пересунути межу поділу двох середовищ - соку та олії - до кінця капіляра. В цьому разі досягається первинний об’єм клітини, а тиск усередині клітини точно зрівноважується тиском, що утворюється в капілярі мікроманометром. Цей тиск, який вимірюється електронним датчиком тиску, відповідає тургорному тиску. Метод характеризується незначною руйнівною дією і дає можливість досліджувати дрібні (до 20 мкм) клітини, вміст рідини в яких становить кілька піколітрів (10-12 л).
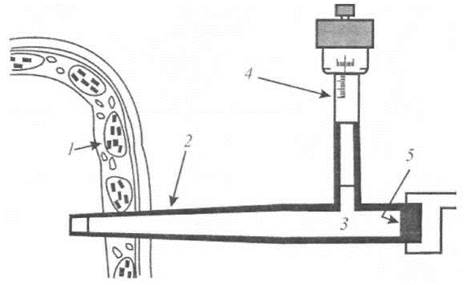
Рис. 6.12. "Зонд тиску"- мікроманометр для вимірювання тургорного тиску в рослинних клітинах [Green and Stanton, 1967]: 1 - Клітина, 2 - мікрокапіляр, 3 - олія, 4 - мікрометр, 5 - датчик тиску.
6.7.7. Аналіз кривих тиск-об’єм
Одним з найкращих є графічний метод вимірювання водного ютенціалу, який базується на побудові та аналізу кривих тиск-об’єм. Ці криві характеризують співвідношення між вмістом води в рослинній тканині та компонентами водного потенціалу. Осмотичний потенціал У тканини пов'язаний з дефіцитом води в ній так:
![]()
де Δв - дефіцит води, ф - осмотичний коефіцієнт, ρ - густина води, R - універсальна газова стала, Т - абсолютна температура, N — загальна кількість молів розчиненої речовини у зразку.
Використовуючи останнє рівняння, можна отримати співвідношення між тиском та об’ємом у вигляді:
![]()
де р - тиск, що відповідає стану рівноваги в камері тиску, V0 - початковий об’єм води, Vв - загальний об’єм виділеної води, Ψm - тургорний потенціал тканини.
Якщо певну кількість води виділити з паростка, тургорний потенціал дорівнюватиме нулю і останнє рівняння набуває такого вигляду:
![]()
Це рівняння характеризує лінійну залежність 1/Ψs від дефіциту води, викликаного лише осмотичним потенціалом.
Процедура вимірювань передбачає зрізування паростка або листка, зважування зразка та розміщення його в камері, де утворено режим насичення. Після остаточної гідратації зразок знову зважують і розміщують у камері тиску з вологою вистилкою. В камері установлюють настільки велику вологість, щоб запобігти випаровуванню з поверхні зразка. Тиск у камері поступово дискретно (з кроком близько 6 атм) збільшують і з кожним кроком вимірюють об’єм виділеної води. Після кожного етапу збільшення тиску і вимірювання об’єму виділеної води оновлюють стан рівноваги у камері тиску. Для цього тиск трохи зменшують перед наступним кроком, чекають, доки настане стан рівноваги і визначають нове значення водного потенціалу зразка. Через 10-15 таких вимірювань зразок прибирають з камери, зважують, висушують і знову зважують. Далі будують криву залежності (1/Ψв) = f(Δc), де Ψв - водний потенціал. Останні точки цієї залежності апроксимують прямою лінією, яка перетинає вісь ординат у точці А, яка відповідає осмотичному потенціалу при повній тургесцентності 1/Ψs100, а вісь абсцис - у точці D, що дає можливість визначити відносний вміст води в апопласті. Криволінійна частина залежності дозволяє оцінити вплив тургорного потенціалу Ψm на повний водний потенціал Ψв (рис. 6.13). Точка, в якій вплив тургорного потенціалу відсутній, відповідає осмотичному потенціалу Ψs0 при витраті тургорного потенціалу (точка В) та величині водного дефіциту при втраті тургорного потенціалу (точка Q. Різниця між криволінійною та прямолінійною ділянками залежності характеризує об’ємний модуль пружності Е. Більш крутій криволінійній ділянці відповідають більш високі значення модуля пружності. Слід відзначити, що модуль пружності зменшується при збільшенні водного дефіциту та зменшенні тургорного потенціалу.
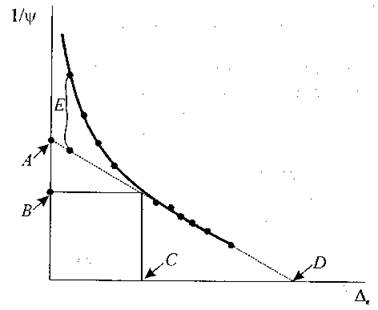
Рис. 6.13. Графічний метод вимірювання водного потенціалу на основі побудови та аналізу кривих тиск-об'єм з метою оцінки співвідношення між вмістом води в рослинній тканині та компонентами водного потенціалу (пояснення в тексті).
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.