БОТАНИКА ТОМ 2 - ФИЗИОЛОГИЯ РАСТЕНИЙ - 2007
6. ФИЗИОЛОГИЯ ОБМЕНА ВЕЩЕСТВ
6.16. Вторичный метаболизм
Процессы, происходящие в организме, которые не относятся к основному обмену веществ — первичному метаболизму, однако являются его следствием, обобщенно называют вторичным метаболизмом, а полученные в ходе этих процессов вещества — вторичными метаболитами. Такие очень разные по химическому строению соединения (уже известно более 200 000 структур) часто встречаются только в определенных группах растений и имеют значение для биохимической систематики. Для каждого вида характерен свой, особый спектр различных вторичных метаболитов, многие из которых синтезируются постоянно, тогда как синтез других индуцируется только после воздействия определенных биотических или абиотических факторов окружающей среды.
Вторичные метаболиты выполняют огромное количество эколого-химических функций (см. гл. 9; 13.8). Они действуют как привлекающие или отпугивающие вещества, препятствуют поеданию растений животными, имеют бактерицидные свойства или являются ингибиторами роста растений-конкурентов (аллелопатия, см. 9.5) Подавляющее большинство вторичных метаболитов формирует своеобразный химический щит, за которым растение успешно скрывается от бесчисленного множества врагов — растительноядных животных и патогенных микроорганизмов (вироидов, вирусов, бактерий, грибов). Если принять во внимание количество потенциальных врагов у растений (две трети всех видов животных являются растительноядными, 30% всех видов грибов, 10—15% всех видов бактерий, 45 % вирусов и все без исключения Вироиды являются фитопатогенными), то в основном неуязвимый или мало уязвимый растительный мир обнаруживает замечательную эффективность защитных мер (количественную), к которым наряду с растительными метаболитами относятся механические барьеры (шипы, колючки, клеточные стенки, кутикула и т.д.). Неудивительно, что довольно много вторичных метаболитов токсичны (табл. 6.24): в целом из растений выделено более 17 000 токсинов, многие из них ядовиты для человека. На протяжении многовековой селекции культурных растений, предназначенных в пищу человеку, их хозяйственная пригодность, как правило, определялась понижением содержания токсинов и горечей, вредных для человека, но необходимых растению в качестве защитных веществ (путем исключения из отбора — элиминации). Как раз поэтому в монокультуры и были отобраны такие относительно беззащитные перед поедающими их животными и патогенами сельскохозяйственные растения, а опасность эпифитотий стала неминуемой. Так, эпифитотия картофеля, вызванная грибом Рhyto-phtora infestans в 1845—1846 гг. в Ирландии стала причиной голода, унесшего жизни почти миллиона человек и вынудившего значительную часть населения (1,5 млн) эмигрировать (в основном в США). Только когда в XX в. удалось в значительной степени заменить естественные защитные механизмы, отсутствующие у культурных растений, химическими средствами защиты растений, снабжение продуктами питания при интенсификации сельского хозяйства достигло высокого уровня.
Таблица 6.24. Основные группы растительных ядов
Класс вещества |
Приблизительное число известных соединений |
Пример |
Источник |
10000 |
Сенеционин |
Senecio jacobaea |
|
Сердечные гликозиды |
200 |
Дигитоксин |
Digitalis purpurea |
Цианогенные гликозиды |
60 |
Амигдалин |
Prunus amygdalus |
Глюкозинолаты |
150 |
Синигрин |
Brassica olerácea |
Фуранокумарины |
400 |
Ксантотоксин |
Pastinaca sativa |
Иридоиды |
250 |
Аукубин |
Aucuba japónica |
Изофлавоноиды |
1000 |
Ротенон |
Dems elhptica |
Непротеиногенные Аминокислоты |
400 |
ß-Цианоаланин |
Vicia sativa |
Полиацетилены |
650 |
Энантотоксин |
Oenanthe crocata |
Хиноны |
800 |
Гиперицин |
Hypericum perforatum |
Сапонины |
600 |
Лемнатоксин |
Phytolacca dodecandra |
Сесквитерпенлактоны |
3 000 |
Гименоксин |
Hymenoxis odorata |
50 |
Вискотоксин |
Viscum album |
|
100 |
Абрин |
Abrus precatonus |
Разностороннее действие, которое оказывают на человеческий организм вторичные метаболиты растений (отравляющее, болеутоляющее, противовоспалительное, дурманящее и т. д.), обнаружилось и стало использоваться уже на очень ранних этапах эволюции человека. Предположительно, фармакология возникла раньше сельского хозяйства. До сих пор вторичные метаболиты представляют собой неисчерпаемый источник лекарственного сырья, например, сердечный гликозид винбластин, применяемый для лечения сердечной недостаточности; таксол — для борьбы с определенными формами рака, отхаркивающее средство кодеин, морфий — в качестве анальгетика.
Далее на примере небольшого числа соединений рассмотрим Строение и Функции некоторых вторичных метаболитов. Из них наиболее богаты представителями фенолы, терпеноиды и алкалоиды.
Фенолы в качестве общего структурного признака имеют как минимум одно ароматическое кольцо, в котором заместителями являются одна или несколько ОН- групп. Последние, со своей стороны, тоже могут подвергнуться замещению (например, —ОСН3, метоксигруппа). К фенолам ведут Различные метаболические пути. Самые значимые из них:
✵ шикиматный путь и метаболические пути, ведущие свое начало от него;
✵ ацетатно-малонатный путь;
✵ путь синтеза терпеноидов (см. 6.12.2);
✵ комбинации всех путей.
Примеры фенолов, образованных в результате шикиматного пути, представлены на рис. 6.114, среди них — переносчики электронов в фотосинтезе, пластохинон и филлохинон (см. рис. 6.56), а также убихинон — компонент окислительновосстановительной системы дыхательной цепи (см. рис. 6.95), α-токоферол, который присутствует в мембранах пластид и защищает Мембранные ЛИПИДЫ от окисления. Юглон представляет собой нафтохинон из плодов и листьев грецкого ореха (Juglans regia), обусловливающий их аллелопатические и антимикробные свойства (см. 9.5, рис. 9.23).
Рис. 6.114. Шикиматный путь синтеза некоторых распространенных групп фенольных соединений. Остаток R представляет все без исключения заместители. Для наглядности происхождение некоторых С-атомов из соответствующих веществ отмечено на рисунке точкой

Важную роль в метаболизме фенолов играет семейство коричных кислот с его многочисленными производными. Малая часть этого семейства представлена на рис. 6.114. Коричная кислота образуется в пластидах из фенилаланина (фенилаланинам- миаклиазная реакция, ФАЛ-реакция, рис. 6.115). ФАЛ — ключевой фермент метаболизма фенилпропанов, который регулируется множеством факторов (например, светом, наличием повреждений, заражением патогенами). Фенилпропаны характеризуются наличием одного бензольного кольца и одной линейной боковой С3-цепочки: например, коричная кислота. Понятие часто используется также для обозначения производных метаболитов этой кислоты. Ингибиторы фермента, такие, как, например, 2-аминоиндан-2-фосфоновая кислота (АИФ), очень помогли при изучении функций ФАЛ и фенилпропанов.
В результате характерных замещений появляются производные коричной кислоты (см. рис. 6.115), образующие вместе с ней семейство коричных кислот. Вариация основной структуры посредством замещения является причиной возникновения множества вторичных метаболитов растений. Вторичные метаболиты — образованные из коричной кислоты кумарины, оказывают отпугивающее действие на растительноядных животных, так как являются горечами (например, присутствуют в доннике, ясменнике). Биосинтез кумаринов представлен на рис. 6.116. Горечи высвобождаются только при повреждении, а в интактной клетке хранятся в вакуолях в виде предшественников.
Рис. 6.115. Синтез и замещение транс-коричной кислоты. 2-Амидоиндан-2-фосфоновая кислота — сильный конкурентный ингибитoр фенилаланинаммиаклиазы

Рис. 6.116. Биосинтез кумарина. Подобным образом реагируют другие представители семейства коричных кислот с образованием соответствующих замещенных кумаринов (см. рис. 6.115)

Посредством β -окисления из коричных кислот образуются фенолкарбоновые кислоты. Из собственно коричной кислоты, таким образом, возникает бензойная кислота, а из нее в результате гидроксилирования в орто-положении — салициловая кислота, фенол с бактерицидным действием; одновременно обсуждается вопрос о ее дополнительном функционировании в качестве сигнального вещества во время индукции системной приобретенной устойчивости (SAR1, см. 9.3.1, 9.3.4). Коричные спирты возникают из коричных кислот в результате восстановления и представляют собой мономерные структурные элементы лигнина. Биосинтез лигнина рассматривается в разделе 6.17.2.
1 От англ. Systemic Acquired Resistance. — Примеч. ред.
Флавоноиды и их производные (основная структура показана на рис. 6.114, последующие структуры — на рис. 6.117) представляют обширную группу вторичных метаболитов с разнообразными функциями. Они в основном присущи покрытосеменным растениям и до сих пор не обнаружены у водорослей, грибов, печеночников и листостебельных мхов. По структуре гетероцикла, содержащего кислород, флавоноиды подразделяются на различные группы, биосинтетические связи которых представлены на рис. 6.117. Общим для всех является остаток флаванового основания. Биосинтез начинается с «активированной» пара-кумаровой кислоты — пара-кумароилкоэнзима А. К этой начальной молекуле с помощью фермента халконсинтазы последовательно присоединяются три молекулы малонил-КоА с последующим карбок- силированием и отщеплением коэнзима А. Так образуется промежуточное соединение кетонной природы, которое циклизуется в халкон с отщеплением четвертой молекулы коэнзима А. Поэтапно, путем образования флаванона, дигидрофлаванола и флаван-3,4-диола, наконец, образуется антоцианидиновая группа, в которой через ненасыщенный гетероцикл конъюгируют между собой п-электронные системы ароматических колец А и Б. Поэтому антоцианидины поглощают видимый свет. Их растворы в зависимости от вида заместителя окрашиваются в цвета от нежнорозового до темно-синего, остальные представленные здесь группы флавоноидов поглощают ультрафиолетовый свет.
Рис. 6.117. Биосинтез некоторых групп флавоноидов из пара-кумароил-КоА и малонил-КоА. Под названиями групп флавоноидов в скобках указаны названия соответственных веществ. Флавоноиды, как правило, запасаются в вакуолях в виде гликозидов (см. текст). Дальнейшие замещения В-кольце (-ОН-, -ОСН3-группы) происходят на этапе образования различных групп флавоноидов. Наряду с этим вместо биосинтеза пара-кумароил-КоА может происходить синтез одной из высоко- замещенных коричных кислот (но не собственно коричной кислоты). Флаванон-З-гидроксилаза и антоцианидинсинтаза относятся к группе Fе2+- и аскорбатзависимых диоксигеназ, которые в качестве дополнительного субстрата окисляют 2-оксоглутарат. Химизм реакций см. на рис. 7.53

Основные структуры флавоноидов варьируют в зависимости от заместителей В-кольце (примеры замещения см. на рис. 6.115), а также в зависимости от гликози- лирования различных позиций (ОН-групп А-кольца и гетероцикла, реже — В-кольца), т.е. здесь наблюдается значительное многообразие структур. Гликозиды флаво-
ноидов запасаются в вакуолях. Они играют роль пигментов, защищающих Клетки от УФ-излучения (высокие концентрации в клетках эпидермиса!), антоцианы (гликозиды антоцианидинов) являются вакуолярными водорастворимыми пигментами в цветках (например, розах, живокости, куколе посевном, бегонии), листьях (в краснокочанной капусте!) и плодах (например, в яблоках), реже — в корнях (бальзамин). Их цвета охватывают все оттенки от бледно-розового до темно-синего и фиолетового, в зависимости от вида заместителя, уровня pH и взаимодействия катионов в содержимом вакуолей. Некоторые антоцианы образуют высокомолекулярные агрегаты с включением ионов металлов, сахаров и других метаболитов. Флавоноиды также, вероятно, обладают свойствами защиты от окисления. Флавоноиды катехолового типа (с двумя соседними ОН-группами В-кольце) выделяются из корней в виде сидерофоров. Бактерии родов Rhizobium (симбионт корневых клубеньков) и Agrobacterium (возбудитель опухоли корончатого галла) используют выделяемые корнями флавоноиды как сигнальные вещества для распознавания растений-хозяев (см. 9.2.1, бокс 9.2).
Путем изомеризации В-кольца из флавоноидов образуются изофлавоны (рис. 6.118). Изофлавон генистеин, выделенный из Genista tinctoria и соевых бобов, является ингибитором тирозинкиназы, который используется при лечении лейкемии. Относящийся к группе изофлавонов даидцеин представляет собой предшественник птерокарпанов (например, глиоцеоллина соевых бобов, Glycine max). При этом речь идет о фитоалексинах. В целом они являются антимикробными вторичными метаболитами, которые синтезируются растением в ответ на патогенное заражение (т.е. индуцирование патогенами — см. 9.3.4). Птерокарпаны и изофлавоны, которые в основном содержатся в бобовых растениях, обладают фунгицидным и бактерицидным действием.
Рис. 6.118. Происхождение изофлавонов в результате окислительной изомеризации флаванонов и образование птерокарпанов из изофлавонов (серым цветом обозначены атомы, принадлежащие изофлавоновым предшественникам)

Наряду с щикиматным путем синтеза А-кольца флавановых производных мы ознакомились со второй возможностью биосинтеза ароматического кольца в растительной клетке, который называется ацетатно-малонатным путем (малойил-КоА образуется в результате карбоксилирования ацетил-КоА, см. рис. 6.102). Поскольку при многократной конденсации ацетатных единиц в этих реакциях (в отличие от биосинтеза жирных кислот) восстановление не происходит, появляются промежуточные соединения, которые не встречаются в свободном виде и называются поликетидами. Они циклизуются в гидроксилированные бензольные кольца. Подобные вещества, возникшие в результате поликетидарома- тизации, называются ацетогенинами. Биосинтетический путь у растений и микроорганизмов, а особенно у грибов и бактерий ведет к синтезу многочисленных производных бензойной кислоты, например, антрахинонов, различных антибиотиков (например, тетрациклинов у стрептомицетов или гризеофульвина у видов Реnicillum), а также различных лишайниковых кислот. Поликетидсинтазы являются многофункциональными ферментами, которые уже производятся рекомбинантным способом и используются в культуре Streptomyces для биотехнологического синтеза антибиотиков. Пример простого биосинтеза ацетогенина представлен на рис. 6.119. Плюмбагин, или нафтохинон, в больших количествах содержат листья представителя росянковых Drosophyllum lusitanicum. Он обладает бактерицидным действием и предположительно защищает ослизненные листовые Органы этих насекомоядных растений от грибковых и бактериальных заболеваний. Следует обратить внимание на то, что очень близкое по строению к плюмба- гину вещество юглон синтезируется совершенно другим способом (см. рис. 6.114): структурное сходство не всегда является показателем биосинтетического родства!
Рис. 6.119. Биосинтез плюмбагина Drosophyllum lusitanicum по ацетатно-малонатному пути. Промежуточное поликето-соединение не присутствует в виде свободного продукта реакции

Третью возможность синтеза ароматических колец предоставляет путь синтеза терпеноидов, рассмотренный в следующем разделе (см. 6.16.2).
Терпеноидами (или изопреноидами) называют все соединения, которые формально можно разложить на отдельные изопреновые составляющие и биосинтез которых начинается с изопентенилпирофосфата (рис. 6.120). Согласно количеству структурных С5-элементов, терпеноиды объединяются в грунпы (табл. 6.25), в которых насчитывается множество представителей, выполняющих огромное количество функций, среди прочих также эколого-химических.
Рис. 6.120. Образование предшественника терпеноидов изопентенилпирофосфата по цитоплазматическому ацетатмевалонатному и пластидному 1 -дезокси-D-ксилулозо-5-фосфатному путям. О распространении того и другого путей среди низших растений и прокариот см. текст

Недавно было выяснено, что растения могут синтезировать С5-единицу — изопентенилпирофосфат — двумя способами (см. рис. 6.120). Таким образом, существуют:
✵ цитоплазматический биосинтез, который начинается с ацетил-КоА, промежуточное соединение — мевалоновая кислота;
✵ пластидный биосинтез, начинающийся с пирувата и D-3-фосфоглицеринового альдегида, — промежуточное соединение 1-дезокси-D-ксилулозо-5-фосфат.
Дезокси-D-ксилуозо-5-фосфатный путь обнаружен у цианобактерий и некоторых других бактерий, тогда как остальные бактерии используют ацетатмевалонатный путь. У зеленых водорослей, вероятно, функционирует только дезокси-D-ксилулозо-5-фосфатный путь, хотя у Euglena gracilis как цитоплазматические, так и пластидные изопреноиды синтезируются по ацетатмевалонатному пути.
Происхождение различных классов терпеноидов высших растений по тому и другому пути — в той мере, в какой оно изучено, — представлено на рис. 6.121; рис. 6.122 наглядно показывает принцип линейной конденсации С5-единиц и биосинтез три- и тетратерпеновых предшественников.
Рис. 6.121. Компартментация биосинтеза терпеноидов высших растений. Еще не ясно, в каком количестве экспортируется в цитоплазму синтезированный изопентенилпирофосфат, ДМАПФ — диметилаллилпирофосфат, ФПФ — фарнезилпирофосфат, ГГПФ — геранилгеранилпирофосфат, ГПФ — геранилпирофосфат, ИПФ — изопентенилпирофосфат

Изопентенилпирофосфат находится в равновесной реакции с изомерной формой диметил аллил пирофосфатом. Монотерпены (С10) появляются в результате присоединения возникшего из диметилаллилпирофосфата после ферментного разрушения С—О-связи карбокатиона к С1 изопентенидпирофосфата (ИПФ) (присоединение мономера к растущей цепи по тину «ГОЛОВА к хвосту») Подобным образом появляются сесквитерпены (С15) из геранилпирофосфата и ИПФ и дитерпены (С20) из фарнезилпирофосфата и ИПФ (рис 6 122) Катализирующие эти реакции ФЕРМЕНТЫ называются пренилтрансферазами. Линейные молекулы геранилпирофосфата (С10), фарнезилпирофосфата (С15) и геранилге- ранилпирофосфата (С20) являются исходными для разнообразных преобразований молекул ряда моно-, сескви- и дитерпенов (см табл 6 25, рис 6 122, 6 123) Только представителей семейства Аsterасеае В настоящее время обнаружено около 1 000 сескви- и дитерпенов.
Рис. 6.122. Модульный принцип синтеза терпеноидов. В результате соединения по типу «голова к хвосту» на предварительном этапе появляются предшественники моно-, сескви- и дитерпенов (а также не изображенных здесь олиго- и политерпенов); в процессе соединения по типу «хвост кхво- сту» двух молекул фарнезилпирофосфата появляется С30-предшественник тритерпенов сквален, а в результате подобного же объединения двух молекул геранилпирофосфата — С40-предшественник тетратерпенов фитоен

Таблица 6.25. Обзор классов терпепов и некоторые типичные представители
Количество С5-единиц |
Класс |
Пример |
Функция(и) веществ(а) |
1 |
Гемитерпены |
Изопрен |
Защита мембраны от высоких температур (?) |
Остаток пренила в цитокининах |
Фитогормоны |
||
Остаток пренила в птерокарпанах |
Фитоалексины |
||
2 |
Монотерпены |
Тимол, ментол, камфара |
Вещества, отпугивающие членистоногих1 |
1,8-Цинеол |
Аллелопатик |
||
3 |
Сесквитерпены |
Сиренин |
Аттрактант гамет Аllomyces |
Капсидол |
Фитоалексин |
||
4 |
Дитерпены |
Фитол |
Закрепление молекул хлорофилла в белке |
Гиббереллины |
Фитогормоны |
||
Таксол |
Фунгицид, ингибитор деления клеток |
||
6 (2 раза по 3) |
Тритерпены |
Фитостеролы (например, ситостерол) |
Структурные элементы мембран |
Сердечные гликозиды (карденолиды) |
Яды, действующие на Сердце и нервную систему |
||
Сапонины (например, дигитонин) |
Бактерицидные вещества с детергентным действием |
||
Брассиностероиды |
Регуляторы роста |
||
8 (2 раза по 4) |
Тетратерпены |
Каротиноиды (каротины, ксантофиллы) |
Дополнительные фотосинтетические пигменты, придающие окраску пигменты |
6-10 |
Олиготерпены |
Прениловые остатки пластохинона, убихинона |
Закрепление в мембране тилакоидов или митохондрий окислительно-восстановительных систем2 |
15 |
Олиготерпены |
Долихол |
Акцептор олигосахаридов для биосинтеза гликопротеинов, закрепляется в ЭР |
>500 - 100 |
Политерпены |
Каучук (полностью цис-изомер) |
Вещество, защищающее растение от поедания животными (в млечном соке) |
Гуттаперча (полностью транс-изомер) |
Вещество, защищающее растение от поедания животными (в млечном соке) |
||
Спорополленины |
Структурный полимер пыльцевой экзины |
1 В низких концентрациях монотерпены привлекают опылителей и являются основными компонентами эфирных масел (например, розы, герани и др.), придают растениям аромат, обладают бактерицидными свойствами. — Примеч. ред.
2 Эту же функцию выполняет фитол хлорофилла, состоящий из 4 изопреновых единиц. — Примеч. ред.
Гемитерпены, для которых типично присоединение к нетерпеноидным молекулам одного или нескольких прениловых остатков, являются птерокарпанами (глицеоллин, см рис 6 118) и цитокининами (см 7 6 2), группы фитогормонов. В сильную жару некоторые растения, прежде всего деревья (в основном виды Quercus и Popuius), синтезируют из диметилаллилпирофосфата изопрен, который выделяется в атмосферу. Вполне возможно (хотя это еще не доказано), что изопрен защищает от теплового повреждения фотосинтетические мембраны. Выработка изопрена может быть значительной и в итоге приводить к 15 — 50% потерям фиксированного углерода. Голубая дымка над лесами во время зноя является следствием эмиссии изопрена Пиретрины (у видов Chrysanthemum) представляют собой гемитерпены, в которых две С5-единицы соединены между собой и с циклопропаном (см рис 6 123). Они являются очень сильными естественными инсектицидами, которые также производятся в коммерческих целях.
Рис. 6.123. Примеры структур характерных представителей различных классов терпеноидов

• Монотерпены в большом количестве обнаруживаются в составе эфирных масел и могут выполнять как аттрактивную, так и отпугивающую функцию (последняя прежде всего направлена против членистоногих) Камфара и 1,8- цинеол — компоненты вырабатываемых Salvia leucophylla в калифорнийской чапаррали летучих аллелопатиков, пары которых распространяются на расстояние 1 — 2 м вокруг кустов шалфея и очень сильно сдерживают рост других растений (см. 9.5).
Сесквитерненом является, например, ювабион в древесине бальзамической пихты (Abies balsámica), который вследствие функций, схожих с действием ювенильных гормонов, тормозит развитие насекомых. Еще один сесквитерпен, сиренин, представляет собой аттрактант гамет (гамон) у живущих в воде плесневых грибов Allomyces (см. 8.2.1.1).
Примеры дитерпенов — фитол, который закрепляет хлорофилл в хлорофилсвязывающих белках (см. 6.4.2) и гиббереллиновый класс фитогормонов (см. 7.6.3). Таксол тихоокеанского тиса (Taxus brevifolid) является высокозамещенным дитерпеном, который запасается в коре и, как полагают, оказывает токсическое действие на ГРИБЫ. Таксол связывается с микротрубочками веретена деления (см. бокс 2.2) и препятствует их деполимеризации: это приводит к остановке митоза. На этом механизме основано цитостатическое действие таксола, которое в настоящее время используется в лечении опухолей (эффективен, например, при лечении рака груди).
Тритерпены образуются в результате димеризации по тину «хвост к хвосту» из двух С15-единиц (фарнезилпирофосфатов) (см. рис. 6.122). Синтезированный сквален циклизуется с образованием стеранового скелета и представляет собой исходное вещество для биосинтеза стероидов (например, фитостеролов, сапонинов, брассиностероидов) и других классов терпенов.
К стероидным гликозидам относятся широко распространенные сапонины. Они встречаются, в частности, в оболочке многих семян, в корнях, корневищах и служат для защиты от бактериального заражения. Токсическое действие сапонинов сказывается на мембранах: сапонины являются детергентами (отсюда и название!)1. Диосгенин, агликон диосгенингликозида (остаток гликозида, который не содержит сахар), на протяжении многих лет вырабатывался из корневищ центральноамериканских тропических лиан рода Dioscorea и использовался в фармацевтике в качестве предшественника полусинтетических стероидов (например, кортикостероидов, ингибиторов овуляции). Однако вскоре исчезновение дикорастущих растений привело к запрету их экспорта. В настоящее время в качестве его предшественников используют желчные кислоты животных, которые поставляются со скотобоен2.
1 От греч. sapo — мыло. — Примеч. ред.
2 В нашей стране разработаны методы биотехнологического получения стероидных гликозидов из культуры клеток Dioсогеа — Примеч. ред.
Определенные штаммы гриба Gaeumanno- myces graminis способны инфицировать корни Avena sativa, после того как разрушат при помощи своего фермента находящийся в эпидермисе корней сапонин (авенацин А-1). Штаммы, не имеющие соответствующего фермента, могут поражать только те виды Avena, которые не содержат сапонин (например, Avena longi- glumis).
К стероидам относятся также сердечные средства — карде нолиды (сердечные гликозиды), например, строфантин и гликозиды наперстянки, в частности, дигитоксин и дигоксин. Последние в основном используются для лечения сердечной недостаточности. В точно отмеренных дозах они замедляют сердцебиение, однако в высоких концентрациях очень ядовиты для млекопитающих. Токсическое действие сердечных гликозидов основано на нарушении возбудимости нервной системы (ингибирование Na+/ К+-АТФазы). Гусеницы бабочки монарха (Dañaus plexippus) живут на ваточнике Asclepias curassavica и запасают сердечные гликозиды в брюшке. Благодаря этому взрослые бабочки становятся несъедобными для своих главных врагов — птиц, так как сердечные гликозиды вызывают сильную дурноту. Молодые птицы быстро приучаются не потреблять бабочку монарха в пищу. Этому способствует бросающаяся в глаза предупредительная окраска бабочки.
Подобно тритерпенам в результате димеризации по типу «хвост к хвосту» из двух С20-единиц с образованием фитоена возникают тетратерпены (геранилгеранилпирофосфат) (см. рис. 6.122). Фитоен является предварительной стадией для биосинтеза каротиноидов (см. рис. 6.50), о которых мы уже упоминали как о дополнительных фотосинтетических пигментах. Каротиноиды придают окраску цветкам (например, виолаксантин у Viola)1 и плодам (красный пигмент томатов ликопин является нециклическим каротиноидом). Каротиноиды обнаружены и в других органах: например, β-каротин в корнеплодах моркови, Dauern carota. Эти плазмохромные (связанные с мембраной) пигменты накапливаются в пластидах (хлоропластах, хромопластах).
1 Окраска, обусловленная каротиноидами, у фиалок встречается крайне редко. Типичную фиолетовую окраску придают антоцианы. — Примеч. ред.
Олиготерпены состоят из 5—15 С5-единиц. Они обнаружены в качестве липофильных группировок в мембранных липидах, например, в убихиноне, пластохиноне, филлохиноне (см. рис. 6.56; 6.95). Долихол- пирофосфат (С75) является донором олигосахаридов в биосинтезе гликопротеинов в эндоплазматическом ретикулуме.
Политерпены каучук (который, например, присутствует в млечном соке Hevea brasïliensis и Parthenium argentatum) и гуттаперча (из Palaquium balata, Sapotaceae) также появляются в результате последовательной конденсации С5-единиц, в каучуке их до 5 000. Полимеры, присутствующие в млечном соке, защищают растения от поедания животными. Каучук (полностью цис-изомер полиизопрена) используется для производства натуральной резины, гуттаперча (полностью трансизомер полиизопрена) ранее использовалась для изоляции электрических проводников. Подобное строение, по-видимому, имеет спорополленин экзины пыльцевых зерен (см. рис. 11.174). Чикл, похожий на каучук политерпен саподиллы, используется в изготовлении натуральной жевательной резинки.
К этой, наверное, самой многочисленной группе вторичных метаболитов в настоящее время относится уже 10000 известных веществ (см. табл. 6.24) с очень разной и иногда очень сложной структурой, содержащихся в низших (грибах!) и высших растениях (рис. 6.124). Богаты алкалоидами, например, пасленовые, маковые, лютиковые и кутровые, бедны ими в основном растения, содержащие большое количество терпенов, например, губоцветные и сложноцветные.
Рис. 6.124. Примеры структур типичных представителей алкалоидов
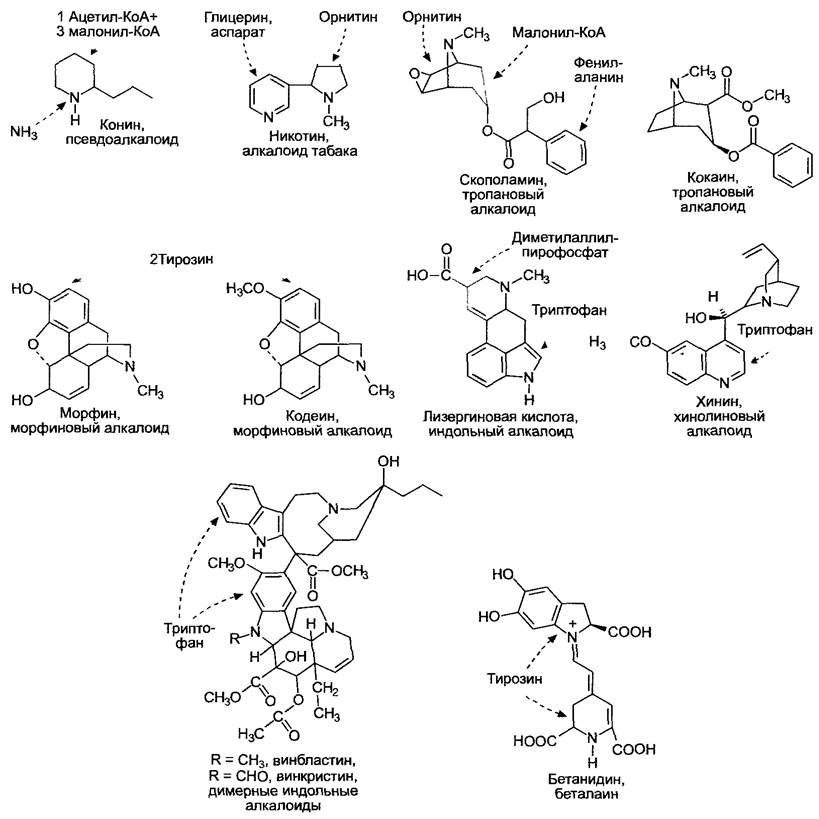
В группу собственно алкалоидов объединяют все вещества, которые содержат гетероциклически связанный азот (поэтому они характеризуются щелочной реакцией); их биосинтез начинается с аминокислот. Алкалоиды часто оказывают на нервную систему позвоночных специфическое действие. Настоящим алкалоидам противопоставляют псевдоалкалоиды,азот которых поставляется не аминокислотой (пример: кониин, яд болиголова, Соnium maculatum, см. рис. 6.124, источником его азота является аммиак). Протоалкалоидаминазывают алкалоиды, образованные из аминокислот, азот которых не включен в гетероцикл (пример: мескалин Lophophora williamsii, рис. 6.125).
Рис. 6.125. Биосинтез мескалина из L-тирозина

Алкалоиды в большинстве случаев являются горечами или токсинами, которые служат растению для защиты от поедания животными; некоторые усиленно синтезируются в процессе заражения патогенами и соответственно представляют собой бактерицидные и фунгицидные фитоалексины (например, бензофенантридиновый алкалоид макарпин калифорнийской эшшольции, Еschscholzia californica). Защитное действие алкалоидов эффективно не только против позвоночных, но и против беспозвоночных животных: никотин табака (Nicotianaа tabacum) является сильнодействующим инсектицидом.
Беталаины, причисляемые к алкалоидам, — это водорастворимые пигменты, к которым относятся желтые бетаксантины и бетацианины (от красных до фиолетовых). Они обнаружены в качестве пигментов цветка у гвоздичных (например, кактусовых, амарантовых, бывшего семейства Сhenopodiасеае — см. 11.2). Беталаины никогда не встречаются вместе с антоцианами. Пигмент столовой свеклы (Beta vulgaris) — бетанидин (см. рис. 6.124), беталаин группы бетацианинов. Беталаином является и пигмент шляпки красного мухомора (Amanita muscaria). Биосинтетический путь беталаинов, очевидно, зарождался в эволюционном процессе как минимум два раза.
Воздействие алкалоидов на центральную нервную систему приводит к наркотической зависимости от этих опьяняющих веществ. Сюда относятся морфий мака снотворного (Papaver somniferum), мескалин кактуса пейота, кокаин коки (Erythroxylum соса) и алкалоиды лизергиновой кислоты спорыньи (Claviceps purpurea), которые были известны еще в античные времена и играли определенную роль в культе богини Деметры. Содержащийся в некоторых пасленовых тропановый алкалоид скополамин был основным действующим веществом мазей средневековых ведьм и в больших дозах вызывал видения с ощущением полета.
Однако многие алкалоиды — это скорее благословение, чем проклятье: они незаменимы в качестве лекарственных средств. Таковы димерные индоловые алкалоиды винбластин и винкристин мадагаскарского катарантуса (Catharanthus roseus), которые используются при лечении лейкемии, хинин хинного дерева (Cinchona), используемый для профилактики малярии, или похожее на морфин вещество кодеин мака снотворного — эффективное отхаркивающее средство.
Мы не можем представить здесь полностью очень сложный биосинтез всех типов алкалоидов, поэтому в качестве примера простого синтеза на рис. 6.125 показано образование протоалкалоида мескалина.
6.16.4. Глюкозинолаты и цианогенные гликозиды
Благодаря тому, что цианогенные гликозиды и глюкозинолаты широко распространены, они являются важными вторичными метаболитами, защищающими растения от поедания животными (кроме того, они исключают присутствие друг друга). Известно около 60 различных цианогенных гликозидов и 150 различных глюкозинолатов. Описано более 2 500 цианогенных видов растений, принадлежащих к разным семействам. Глюкозинолаты встречаются в основном в семействах порядка Capparales
(например, Вгassicасеае, Сарраridасеае, Тгораeolасеае). Резуховидка Таля (Arabidopsis thaliana из сем. Вrassicасеае) содержит более 25 различных глюкозинолатов.
Глюкозинолаты и цианогенные гликозиды образуются из аминокислот и на первых этапах синтезируются одинаково (синтез промежуточного вещества альдоксима, рис. 6.126). В дальнейшем сходство между ними наблюдается в процессе запасания конечных продуктов реакции в форме гликозидов в вакуолях, где они накапливаются в высоких концентрациях в качестве предшественников веществ, предохраняющих растение от патогенов и поедания животными. После разрушения Ткани гликозиды расщепляются ферментами, которые в интактной клетке отделены мембранами от своего субстрата (рис. 6.127).
Рис. 6.126. Биосинтез цианогенных гликозидов и глюкозинолатов, их примеры. Хотя процессы биосинтеза протекают на первых этапах одинаково до образования альдо- ксима и оба класса веществ ведут свое начало от аминокислот, цианогенные гликозиды и глюкозинолаты ни разу не были обнаружены вместе. Представленные здесь цианогеные гликозиды содержат в качестве сахарного компонента глюкозу. Кроме глюкозы встречаются и другие сахара, например, гентиобиоза в амигдалине (агликон, такой, как пруназин).

Рис. 6.127. Запасание и расщепление цианогенных гликозидов и глюкозинолатов. Цианогенные гликозиды (А) и глкжозинолаты (В) представляют собой предшественников токсинов, из которых основной токсин (синильная кислота (изотиоцианат), нитрил) образуется только после разрушения клеточной структуры, например, в результате поедания животным. В интактных клетках субстраты и ферменты разделены благодаря компартментации. В случае дуррина проса (Sorghum bicolor) цианогенный гликозид накапливается в вакуолях клеток эпидермиса, ß-глюкозидаза сохраняется в хлоропластах, а гидроксиллиаза — в цитоплазме подлежащего мезофилла. Глюкозинолат- синигрин хрена деревенского (Armoracia rusticana) находится в вакуолях, мирозиназа — в цитоплазме тех же клеток

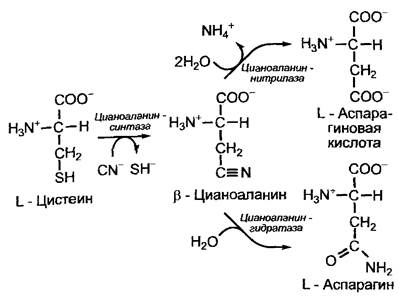
Из цианогенных гликозидов наряду с сахаром (часто глюкозой или гентиобиозой) образуется циангидрин, который расщепляется гидроксинитриллиазами на альдегид и синильную кислоту (НСN). Синильная кислота представляет собой сильный ингибитор цитохромоксидазы, нарушающий митохондриальное Дыхание (см. 6.10.3.3). Растения обезвреживают синильную кислоту, которая, кроме того, постоянно образуется в небольших количествах во время биосинтеза этилена при участии β-цианоаланинсинтазы и в процессе преобразования β-цианоаланина в аспарагин и аспарагиновую кислоту (рис. 6.128).
Рис. 6.128. Обезвреживание цианида (CN-) высшими растениями
Ферментативное расщепление глюкозинолатов мирозиназой наряду с глюкозой дает нестабильный агликон, который распадается на различные продукты, в основном изотиоцианаты (горчичные масла) и нитрилы, их образование также контролируется ферментами (см. рис. 6.127). Горчичные масла обладают резким запахом и острым вкусом (хрен деревенский!) и раздражают слизистые оболочки. Горчичные масла разрушают мембраны, оказывая, таким образом, токсическое действие на бактерии и грибы. Судьба тиоцианатов в растении неизвестна. Синтезированные нитрилы разлагаются нитрилазами на аммиак и соответствующие карбоновые кислоты. Предполагается, что индолацетонитрил, полученный из глюкобрассицина, по меньшей мере, на определенных этапах развития растения (прорастание) является предшественником гормона роста индолил-3-уксусной кислоты (см. 7.6.1.2).
Как глюкозинолаты, так и цианогенные гликозиды постоянно синтезируются и расщепляются, т. е. их можно рассматривать исключительно в качестве резервных форм защитных веществ. Вполне возможно, что они, как минимум в некоторых определенных ситуациях, представляют собой также своеобразный запас азота и серы (глюкозинолаты), особенно в корнях и семенах, где обнаружены высокие концентрации этих веществ. При прорастании семян, например, содержание глюкозинолатов в них резко снижается.
Достоверным фактом считается то, что вторичные метаболиты растений, помимо всего прочего, представляют собой важный компонент защиты растений против растительноядных животных и патогенов (см. 9.3, 9.4) и что именно большое количество отдельных защитных мер, к которым также причисляют широкий спектр вторичных метаболитов (в растении могут находиться сотни различных веществ), создает широкую эффективную, но неспецифическую защиту1. Правда, в результате химической коэволюции специализированные виды растительноядных животных и патогенов адаптировались к определенным видам растений и начали успешно обходить их защитные химические приспособления.
1 Для обозначения неспецифической защиты к широкому кругу патогенов в русской литературе используют термин «горизонтальная устойчивость». — Примеч. ред.
Иногда они даже используют защитные вещества растений в своих целях. Так, многие глкжозинолаты являются эффективными веществами, отбивающими аппетит у большинства животных, питающихся гусеницами капустницы (Pieris brassicae), которые содержат глкжозинолаты (например, сянегрин). Ранее уже говорилось о том, что гусеницы бабочки монарха накапливают поступающие с пищей сердечные гликозиды (кормовое растение рода Ascle - pías) (см. 6.16.2). Последние передаются имаго и защищают насекомое от врагов — в основном птиц.
Алкалоиды люпина весьма токсичны для большинства животных. Понадобилось много сил и труда селекционеров, чтобы вывести безалкалоидный (сладкий) люпин в качестве кормового растения. В смешанных посевах сладкого люпина и люпина, который содержит алкалоиды, сладкий люпин в течение короткого периода времени исчезает в результате деятельности растительноядных животных, тогда как его ядовитый сородич выживает. На наличии люпиновых алкалоидов (например, спар- теина) основана также токсичность ракитника метельчатого (Cytisusscoparius). Однако тлю Acyrthrosiphon spartii спартеин привлекает, в результате она занимает на своем кормовом растении экологическую нишу, недоступную другим животным.
Особенно хорошо изучена химическая экология пирролизидиновых алкалоидов (рис. 6.129). Эти алкалоиды встречаются у родов из семейства Asteraceae (например, Senecio, Eupatorium), а также у Boraginaceae, урода Crotalaria (бобовые) и рода Phalaenopsis (орхидные), и в отдельных случаях — в других семействах. Пирролизидиновые алкалоиды (например, сенеционин) представлены в растении в качестве полярных, водорастворимых аминоксидов и являются горькими, токсичными, а для насекомых митогенными (индуцирующими деление клетки) веществами, которые защищают растение от поедания животными. При попадании в кишечник аминоксиды восстанавливаются до липофильных, третичных аминов, которые легко диффундируют в клетки, где окисляются под воздействием монооксигеназ, содержащих цитохром Р450, до производных пирролина. Они сильно токсичны для печени и дыхательных путей и представляют собой активные алкилирующие агенты. Личинки бабочки медведицы Thyria jacobaea (Arctiidae) получают пирролизидиновые алкалоиды из своего кормового растения Senecio jacobaea и сохраняют их на протяжении всех стадий метаморфоза; пирролизидиновые алкалоиды из семейства Arctiide, Utetheisa omatrix (они принадлежат кормовому растению рода Crotalaria, бобовые), даже переходят из родительского организма в яйца. Отсутствие токсичного действия этих алкалоидов на адаптировавшиеся виды объясняется тем, что поступившие из кишечника в клетки тела липофильные третичные амины снова окисляются до полярных солеобразующих аминоксидов (см. рис. 6.129). Личинки и имаго Arctiide благодаря накопленным алкалоидам эффективно защищены от своих врагов (например, от муравьев) так же, как и яйца. Яркая предупреждающая окраска гусениц и имаго способствует этому.
Рис. 6.129. Химическая экология пирролизидиновых алкалоидов. После попадания в организм растительноядного животного полярный алкалоид в виде аминоксида восстанавливается в кишечнике, третичный липофильный амин попадает в клетки, где он окисляется до токсичного производного пирролина. Адаптировавшиеся насекомые (например, личинки Arctiidae) обезвреживают третичные амины, превращая их снова в полярные аминоксиды и накапливая в клетках. Некоторые виды насекомых синтезируют из потребленных пирролизидиновых алкалоидов свои мужские половые аттрактанты, например, гидроксиданаидал
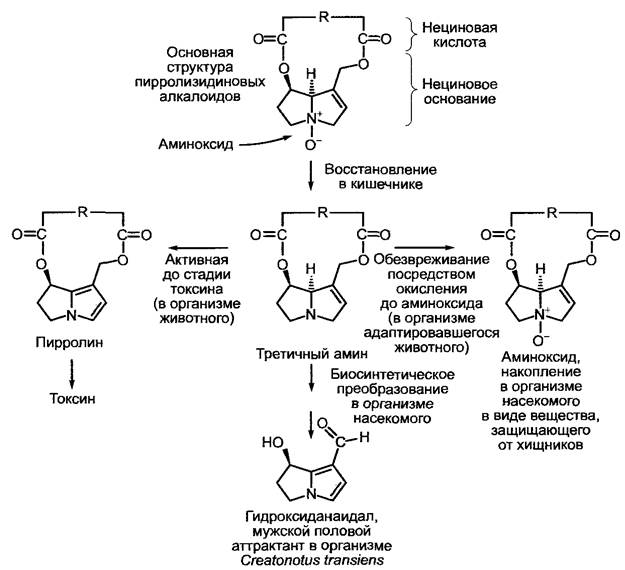
Бабочек подсемейств Danainae (например, виды рода Danaus) и Ithomiinae привлекают пирролизидиновые алкалоиды, их потребляют только взрослые бабочки, например, с нектаром, однако очень часто они экстрагируют алкалоиды из других частей растения, выделяя из хоботка особую жидкость, которую затем всасывают назад уже с растворенными в ней алкалоидами. Растение при этом не служит пищей, поэтому данный случай назвали фармакофагией. Алкалоиды могут составлять 2—20 % сухого вещества животного. Некоторые бабочки, накапливающие пирролизидиновые алкалоиды (например, Danaus plexippus и Аrctiide Сгеatonous transiens), синтезируют из них мужские половые аттрактанты (половые феромоны, например, гидроксиданаидал у Сгеаtonotus).
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.