Фізіологія рослин - Мусієнко М.М. 2001
Фотосинтез: фізіолого-біохімічні та екологічні аспекти
Структурна організація функціональних компонентів електрон-транспортного ланцюга хлоропластів
Існування фотосинтетичної одиниці поставило питання про механізм Міграції енергії до фотохімічних центрів та організацію пігментів у даній системі.
Як відомо, вищі рослини мають три групи пігментів: хлорофіли а, b та каротиноїди. Всі вони зв’язані з білками і складають відповідні комплекси. Так у фотосистемі II основні з них наступні:
СР 47 (СР — від Chlorophyll protein, тобто хлорофіл-білок) з молекулярною масою -50 кД;
СР 43 з масою -43 кД;
світлозбираючий комплекс два (СЗК II), що містить чотири поліпептиди масою 29, 27, 26 та 25 кД.
Кожен із комплексів СР-47 та СР-43 містить до 50 молекул хлорофілу а, тоді як з кожним поліпептидом СЗК II зв’язано відповідно 10-13 молекул хлорофілу а + хлорофілу b. Співвідношення хл а : хл b для комплексу СЗК II дорівнює 1:3.
Щодо фотосистеми І, то в ній знайдено теж три основні комплекси:
СР 62;
СР 58;
світлозбираючий комплекс один (СЗК І), який містить чотири поліпептиди з масою 23, 22, 21 та 20 кД. Зауважимо, що світлозбираючий комплекс І додатково підрозділяється, відповідно на СЗК-la та СЗК-Ів.
З комплексами СР-62 та СР-58 зв’язано близько 50 молекул хлорофілу а, тоді як кожен з комплексів СЗК Іа та СЗК-Ів містять 38 молекул хлорофілу а та 12 молекул хлорофілу в. Хлорофіл а реакційного центра фотосистеми II Р680 та хлорофіл а реакційного центра фотосистеми І Р700, по всій ймовірності, розміщені в комплексах СР-47 та СР-62 відповідно.
Хлорофіл а присутній в кількох формах: в СЗК II — хлорофілу а660, хлорофілу а670, хлорофілу а677 і хлорофілу а684; в комплексах СР43 та СР47 — хлорофілу а660, хлорофілу а670, хлорофілу а677 та хлорофілу а684, і нарешті, в фотосистемі 1 — хлорофілу а662, хлорофілу а668, хлорофілу а679, хлорофілу а687, хлорофілу а695 хлорофілу а705.
Виходячи з уявлення про фотосинтетичну одиницю як ансамбль світлозбираючих молекул пігментів, що обслуговують один РЦ, перенесення енергії можна уявити так:
фотосистема II: СЗК II → СР43→ СР47,
Фотосистема І: СЗК І → CP58→ СР62.
Ефективність перенесення енергії від СЗК II до СР 43 становить майже 100%, як і при перенесенні енергії від хлорофілу в до хлорофілу а, тому що флуоресценція майже ніколи при цьому не спостерігається. В антенному комплексі може бути деяка втрата енергії, оскільки антена флуоресціює. Отже, енергія поглинута різними молекулами антени, мігрує до реакційних центрів, чим і пояснюється той факт, що навіть під впливом слабкого світла майже негайно як результат фотосинтезу починає виділятися кисень. Основним фактором, що визначає ефективність, є час, який потрібний для того, щоб збудження досягало РЦ і розпочався розподіл зарядів:
![]()
Специфікою хлорофілів реакційних центрів є той факт, що вони виступають як димери. Розташування обох молекул хлорофілів в димері відносно біфункціональних лігандів таке, що відстань між сг-орбітами, яка дорівнює 0,36 нм, забезпечує оптимальне перекривання. Віддача електрону збудженим димером проходить легше, ніж збудженим мономером. Проблема розподілу заряду зводиться до проблеми первинного акцептора (а відповідно і донора) електронів в реакційному центрі. Його просторове розміщення повинно бути таким, щоб відновлення акцептора відбулось раніше, ніж проміжний катіон хлорофілу знову прийняв електрон.
За даними Р. Дональда та О. Говінджі (1987), час необхідний для розділення зарядів в РЦ фотосистеми І становить менше 50-80 пс, а для фотосистеми II в межах 150-200 пс, інакше воно не може конкурувати з флуоресценцією. Є дані, що в ФС І час розподілу заряду може бути менший 10 пс.
Розподіл енергії збудження між ФС І та ФС II може змінюватися. Допускають міграцію СЗК II на відносно значні відстані від ФС II до ФС І. Можливо, це пояснюється тим, що енергія, яка поглинається в основному ФС II, сприяє нагромадженню відновленого пластохінону, який активує кіназу, що викликає фосфорилування комплексу СЗК II з наступною міграцією його (рис. 51).
Асиметрія фотосинтетичних мембран створює структурну основу для направленого (векторного) функціонування фотосинтетичної мембрани при перетворенні світлової енергії.
Вважають, що комплекси ФС І та ФС II знаходяться в різних ділянках мембрани (рис. 52). Більшість РЦ фотосистеми II, їх СЗК та компоненти водоокислення локалізовані в зоні міжмембранних контактів, причому їх рухомість обмежена ван-дер-ваальсовими силами притягання.
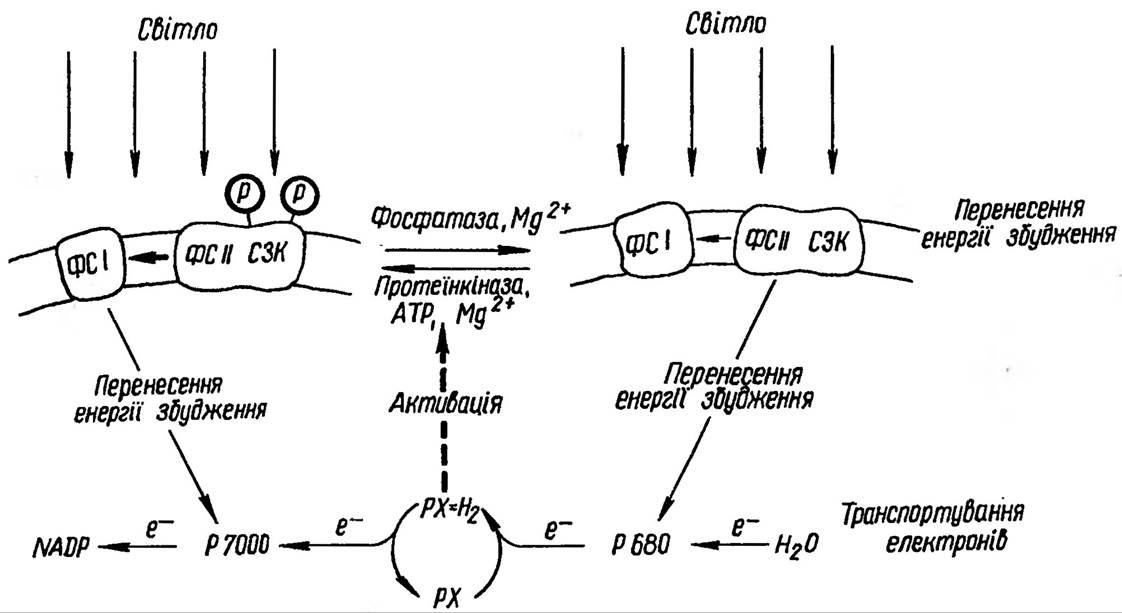
Рис 51. Модель регуляції розподілу енергії електронного збудження при фотосинтезі.
Обернене фосфо- рилування світлозбираючого комплексу в поєднанні з окислювально-відновним станом пластохінону (Рх) забезпечує розподіл енергії між фотосистемою І та фотосистемою II. Відновлення пластохінону фотосистемою II спричинює активування кінази та фосфорилування СЗК ФС II. Внаслідок цього частка енергії електронного збудження, яка передається на фотосистему І, зростає. Окислення пластохінону інактивує кіназу, фосфатаза дефосфорилює СЗК ФС II. Внаслідок цього збільшується частина енергії, що надходить до ФС II. Окислювально-відновний стан пластохінону залежить від розподілу енергії між фотосистемами, тому обернене фосфорилування СЗК ФС II замикає ланцюг оберненого зв'язку. При найменшому порушенні рівноваги система завжди прагне повернутись до вихідного збалансованого стану що діють між поверхнями сусідніх мембран Реакційні центри ФС І та їхні світлозбираючі комплекси (СЗК І) локалізовані переважно в тих частинах мембран, які обернені до строми. Комплекси спрягаючого фактора розподілені в мембрані аналогічно ФС І, а комплекс цитохромів b6—f розміщений, мабуть, досить рівномірно. Він зберігає латеральну рухомість, що дає йому змогу дифундувати в зону міжмембранних контактів і в зворотньому напрямку.
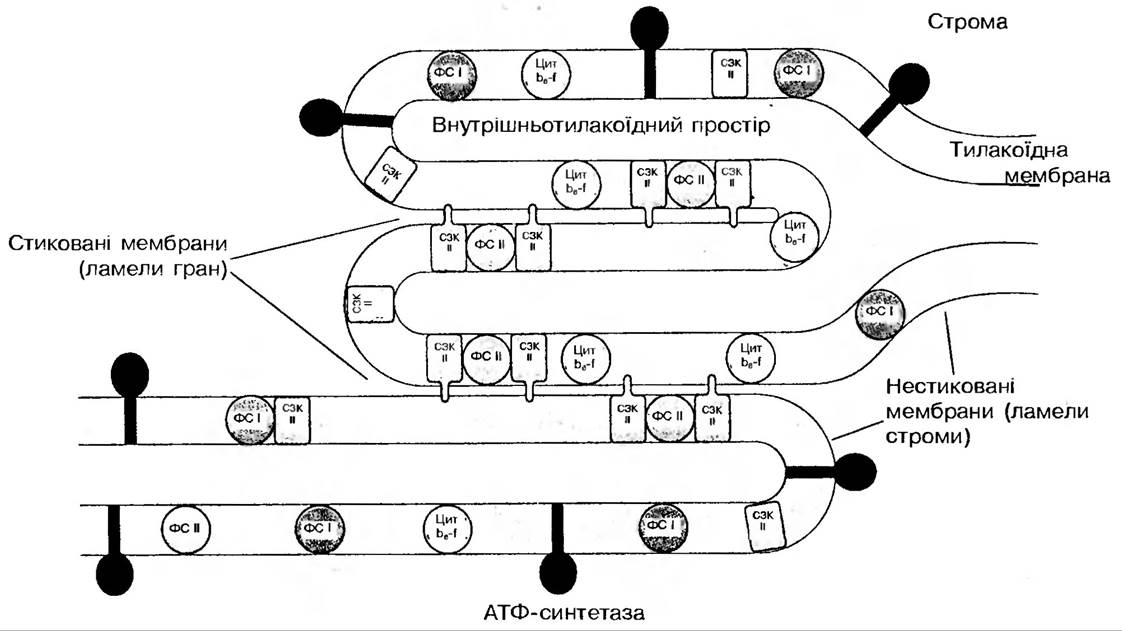
Рис. 52. Розподіл структурно-функціональних комплексів в стикованих і нестикованих ділянках тилакоїдної мембрани
Латеральна асиметрія в розподілі білкових комплексів мембрани визначається, насамперед упаковкою мембран в стопки — грани.
Последнее обновление: 23/05/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.