БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 4. МОЛЕКУЛЯРНА ОРГАНІЗАЦІЯ І БІОЛОГІЧНІ ФУНКЦІЇ МЕМБРАН
4.1. Структура біологічних мембран
Перші прямі хімічні Методи в дослідженні мембран були застосовані Е. Овертоном (1895-1902). Установивши, що в клітину швидше проникають речовини, які розчинні в ліпідах, він запропонував першу модель плазматичної мембрани (1902), а саме: мембрана - це тонкий шар фосфоліпідів. У 1925 р. Е. Гортер і Ф. Грендель показали, що площа моношару ліпідів, екстрагованих з еритроцитів, у два рази більша, ніж сумарна площа еритроцитів. На підставі цих досліджень було зроблено припущення, що Ліпіди в мембрані розміщуються у вигляді бімолекулярного шару товщиною 3-4 нм. Електрична ємність такої структури складає 0,5 · 10-2 Ф/м2, що відповідає товщині діелектрика - ліпідного шару мембрани - 3,5 нм.
Одночасно з цими дослідженнями було встановлено, що значення коефіцієнта поверхневого натягу на межі розподілу "Клітина-Вода" ближчі до аналогічного параметру на межі "білок- вода" (0,1 дин/см), ніж на межі "ліпід-вода" (~ 10 дин/см). Таке протиріччя було усунуто Дж. Даніеллі й Х. Давсоном, які запропонували в 1935 р. "бутербродну" модель будови біологічної мембрани (рис. 4.1, 1). Така мембрана утворена двома шарами білків з ліпідним бішаром між ними. Пізніше (1956) з метою узгодження моделі з відомими на той час факторами полегшеної дифузії через мембрану низькомолекулярних гідрофільних речовин Дж. Даніеллі та В. Стейн припустили існування в мембрані полярних білкових пор (рис. 4.1, 2).
Завдяки прогресу в техніці приготування ультратонких зрізів (1950-1960) тканин для електронної мікроскопії стало відомо, що тришарова структура притаманна всім дослідженим на той час мембранам. Базуючись на цих морфологічних даних, Дж. Роберт- сон у 1960 р. висловив концепцію унітарної мембрани (рис. 4.1, 3). Ця модель подібна до моделі Даніеллі - Давсона, у ній також відображена ще одна особливість мембрани - її асиметрія (на рисунку білкові шари зображені не однаково). Розвиток електронно-мікроскопічної техніки дав можливість "побачити" мембрану не як суцільну тришарову лінію, а як гранулярну структуру, що має розриви (білкові пори) і ніби складається з окремих глобулярних частин. Більш того, при розбиранні мембран за допомогою детергентів (див. далі) отримували ліпопротеїнові фрагменти, які після видалення детергентів знову збиралися в мембраноподібні структури. Ці дані викликали сумніви щодо моделі Робертсона та породили низку моделей, котрі відображали концепцію глобулярної організації мембран. Прикладом такої стала модель Ф. Шестранда (1963), Дж. Лаці (1964), в якій мембрана уявлялась як чергування ліпідних міцел і білкових глобул, Д. Гріна і Дж. Пердью (1966) - модель мембрани мітохондрій із ліпопротеїнових субодиниць складної конфігурації, А. Бенсона (1966) - вуглеводневі ланцюги ліпідів переплетені з поліпептидами й утримуються за рахунок гідрофобних взаємодій. У моделі Дж. Фінеана (1966) підкреслюється різниця в розподілі компонентів мембран і вперше зроблена спроба описати мембрану з урахуванням конкретних ліпідних молекул (рис. 4.1).
Відкриття фазового поліморфізму водно-ліпідних систем і рідинно-кристалічного стану ліпідів за фізіологічних температур, концепція плинності ліпідного бішару, вимірювання швидкості латеральної дифузії в мембранах (1968-1971) стали підставою для С. Сінджера й Г. Ніколсона запропонувати принципово нову модель молекулярної організації мембран (1972), яку назвали рідинно-мозаїчною (рис. 4.1, 10). Основними постулатами такої мембрани є:
✵ мембрана - це "море" ліпідів, в якому "плавають" окремі білки;
✵ мембранних білків є два типи: периферичні (зв'язані з мембраною за рахунок полярних та іонних взаємодій), інтегральні (пронизують ліпідний бішар і взаємодіють з ліпідами своїми гідрофобними ділянками);
✵ молекули мембран перебувають у постійному латеральному русі тощо. На рис. 4.1,10 зображено рідинно-мозаїчну модель плазматичної мембрани з урахуванням зв'язку її структурних компонентів з цитоскелетом і глікокаліксом.
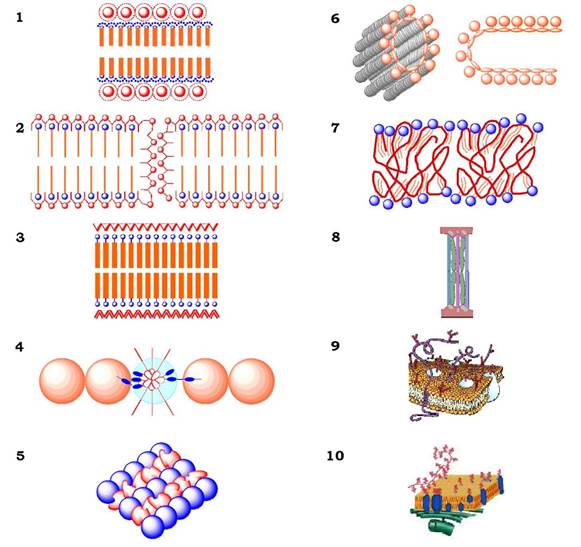
Рис. 4.1. Моделі структури біологічних мембран:
1 - Даніеллі - Давсона; 2 - Стейна - Даніеллі; 3 - Робертсона; 4 - Лаці; 5 - Шестранда; 6 - Гріне й Пердью; 7 - Бенсона; 8 - Фінеана; 9 - Сінджера - Ніколсона (рідинно-мозаїчна); 10 - рідинно-мозаїчна модель з урахуванням зв'язку мембран з цитоскелетом і з глікокаліксом
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.