Молекулярная биология: Структура и функции белков - Степанов В.М. 2005
Посттрансляционная модификация белка
Внутримолекулярные перегруппировки в белках
Полипептидная цепь — первичный продукт биосинтеза белка — часто подвергается химическим превращениям, изменяющим ее ковалентную структуру. Такие превращения могут происходить как в процессе трансляции (котрансляционная модификация), так и после ее окончания, нередко уже после сформирования пространственной структуры белка. Многообразие этих реакций, завершающих созревание белка или видоизменяющих его функциональные свойства, очень велико — их число достигает 300-400. Иногда реакции посттрансляционной модификации называют процессингом (от англ. processing — обработка), подчеркивая этим, что они как бы завершают Биосинтез белка. Этот термин не вполне точен, поскольку некоторые реакции, например фосфорилирование, могут происходить с одной и той же молекулой неоднократно, регулируя ее активность, другие, вероятно, служат сигналом к расщеплению белка.
Обычно реакции посттрансляционной модификации катализируются специфическими ферментами, хотя известны и внутримолекулярные превращения белков, происходящие без участия «внешних» ферментов. Реакции посттрансляционной модификации, как правило, имеющие функциональный смысл, направленность, принято отличать от более или менее случайных повреждающих реакций, также весьма многочисленных, которые происходят с белками in vivo, отражаясь на их структурной целостности и функциональных свойствах. К ним относят, например, реакции окисления, случайный протеолиз и некаталитическое дезамидирование белка, присоединение глюкозы к аминогруппам некоторых белков, в частности гемоглобина, при ее повышенном содержании в крови при диабете и т.п.
Реакции посттрансляционной модификации являются нематричными процессами, они не кодируются непосредственно, поэтому нередко протекают неполно, что приводит к образованию множественных форм того или иного белка — продуктов частичного превращения.
Каков функциональный смысл реакций посттрансляциоиной модификации? На этот вопрос в обобщенной форме можно дать следующий ответ. Во-первых, несмотря на удивительное богатство структур, образуемых аминокислотными остатками и их ансамблями в пространственной структуре белка, это многообразие далеко не безгранично. Разумеется, образование комплексов с кофакторами (металлами, коферментами и т.п) резко расширяет функциональные возможности белка. Однако и этот путь не всегда оказывается достаточным. В таких случаях структурная палитра белка может быть обогащена путем направленного преобразования боковых цепей тех или иных аминокислотных остатков, а иногда и молекулы в целом. Во-вторых, что не менее важно, некоторые реакции посттрансляционной модификации, в особенности обратимые, позволяют управлять активностью белка или целых групп белков в ответ на изменяющиеся потребности Клетки.
Заметим, что реакции посттрансляционной модификации, в отличие от предшествующих: им стадий трансляции, очень индивидуальны, иногда они свойственны только одному белку. К таким реакциям относятся, например, гидроксилирование остатков пролина в коллагене, иодирование тиреоглобулина и некоторые другие. Специфичность посттрансляционной модификации открывает перспективу избирательного воздействия на Процессинг и, следовательно, функцию тех или иных белков.
Далее мы рассмотрим некоторые важнейшие реакции посттрансляционной модификации.
Эта группа превращений стоит особняком, поскольку они не катализируются каким-либо ферментом, а являются результатом внутримолекулярной реакции, направляемой, по-видимому, специфической пространственной структурой белка-предшественника. Последнее обстоятельство сближает такие реакции с каталитическими, поэтому иногда их описывают как автокаталитические процессы, хотя аналогия не вполне справедлива, ибо в этом случае «Активный центр» действует не многократно, как у ферментов, а только однажды.
Профермент гистидиндекарбоксилазы молочнокислых бактерий Lactobacillus, а также Clostridium perfringens синтезируется в виде одной полипептидной π-цепи с молекулярной массой 34 кДа. При внутримолекулярной активации она превращается в цепи а и ß с молекулярными массами соответственно 25 и 9 к Да. Цепь а несет на аминном конце остаток пировиноградной кислоты, который выполняет в активном центре фермента роль своеобразного кофактора и взаимодействует с аминогруппой гистидина, подвергающегося декарбоксилированию. Этот пирувоильный остаток функционально аналогичен пиридоксальфосфату — типичному кофактору ферментов аминокислотного обмена. Заметим, что π-цепь и продукт ее расщепления, построенный из а- и ß-цепей, образуют гексамерную четвертичную структуру.
Особенно интересен путь расщепления я-цепи на а- и ß-цепи, не являющийся гидролитическим. В последовательности аминокислот, которая соединяет будущие а- и ß-цепи,

по-видимому, под влиянием пространственно сближенных функциональных групп прогистидиндекарбоксилазы происходит такая перегруппировка, в которой непосредственно участвуют соседствующие остатки серина. Сначала карбонильная группа предшествующего остатка серина переносится с аминной на гидроксильную группу следующего остатка:
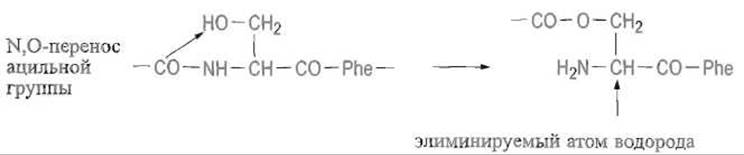
Это так называемая N, О-ацильная миграция. Затем ацильный компонент (ß-цепь) негидролитически элиминируется за счет отщепления а-атома водорода остатка серина (ß-элиминирование), оставляя на аминном конце уже образовавшейся a-цепи весьма неустойчивый остаток дегидроаланина:
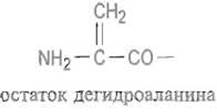
Дегидроатанин, по-видимому, изомеризуется в иминопроизводное пирувоильного остатка, которое затем реагирует с водой, отщепляя аммиак и превращаясь в пирувоильный остаток:
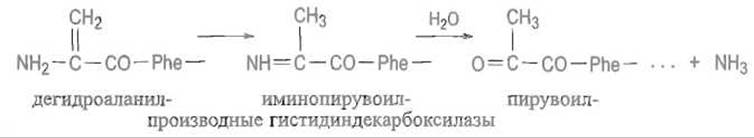
Следует подчеркнуть, что собственно расщепление полипептидной цепи происходит в этом случае без участия воды, негидролитическим путем. Интересно, что у других ферментов аминокислотного обмена — лиаз, отщепляющих аммиак от фенилаланина или гистидина с образованием соответствующих непредельных кислот (коричной в случае фенилаланина или урокаиновой в случае гистидина), наблюдается похожее превращение, которое приводит, однако, не к расщеплению пептидной цепи, а к образованию дегидроаланина, по-видимому, стабилизированного в пространственной структуре фермента. Вероятно, предшественником дегидроаланина в этих случаях является активированное производное серина или цистеина. Дегидроаланин в таких ферментах входит в состав каталитического центра, выступая в качестве электрофильной группы. Заметим, что электрофильные группы не свойственны традиционно построенным белкам.
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.