Основы молекулярной биологии - В.И. Резяпкин 2009
Организация гемонов
Транскрипция геномов ДНК-содержащих вирусов
Молекулярный механизм транскрипции у вирусов принципиально сходен с таковым клеточным.
Особый интерес у вирусов представляет регуляция транскрипции. Это связано с тем, что различные Белки должны синтезироваться в разных количествах: белки вириона - в больших, ФЕРМЕНТЫ - в меньших. А также и с тем, что гены вирусов экспрессируются с различной интенсивностью в процессе их жизненного цикла.
Регуляция транскрипции в значительной степени осуществляется посредством промоторов и терминаторов. Кроме того, первичные транскрипты могут подвергаться процессингу. Для иРНК вирусов эукариот характерны сплайсинг, кэпирование 5’-конца и полиаденилирование 3’-конца. Следует отметить, у вирусов эукариот и фагов отличается процесс трансляции. Прежде всего, это связано с тем, что иРНК фагов может быть полицистронной, а иРНК вирусов эукариот, как правило, функционально - моноцистронна.
Транскрипция геномов ДНК-содержащих фагов
Транскрипция генов ДНК-содержащих фагов может осуществляться как клеточными РНК-полимеразами, так и вирусспецифическими. Например, транскрипция генома нитчатых фагов (М13, fd и др.) осуществляется клеточной РНК-полимеразой. В их геноме закодировано 10 белков. Гены этих фагов можно разделить на две группы: активно транскрибируемые гены и гены, транскрибируемые с меньшей скоростью. Продукты генов первой группы требуются в значительных количествах. Область генома, включающая эти гены, имеет несколько сильных промоторов и сильный терминатор. В связи с этим транскрипция генов, расположенных в этой области, осуществляется с различных промоторов, в то время как терминация происходит только в одном участке. И тогда в результате транскрипции образуются иРНК различной длины с одинаковыми 3’-областями. Белки, закодированные со стороны 3’-конца иРНК, будут, в конечном счете, представлены большим числом цистронов (рис. 11.11).

Рис. 11. 11. Активно транскрибируемые гены нитчатых фагов
Гены второй группы кодируют минорные белки, необходимые в незначительных количествах. Их промоторы являются слабыми, что определяет низкую эффективность транскрипции.
Таким образом, различный уровень экспрессии генов у нитчатых фагов определяется силой промоторов и особенностью синтеза иРНК, отличающихся друг от друга 5’-областями.
У фага SPO1, развивающегося в клетках B.subtilis, регуляция транскрипции осуществляется иным способом. В его геноме выделяют три группы генов: ранние, средние и поздние. Ранние гены транскрибируются сразу же после инфекции в течение 4-5 мин, затем транскрибируются средние гены, через 8-12 мин транскрипция средних генов замедляется, и начинают транскрибироваться поздние гены. Ранние гены фага не отличаются от клеточных генов и транскрибируются клеточной РНК-полимеразой, субъединичный состав которой a2ßß’σ55. σ55- фактор определяет способность РНК-полимеразы узнавать промоторы. Одним из продуктов ранних генов является белок gp28. Этот белок заменяет в составе РНК-полимеразы σ55-фактор и присоединяется к минимальному ферменту a2ßß’, образуя комплекс a2ßß’gp28. В таком состоянии РНК-полимераза теряет сродство к промоторам ранних генов и начинает транскрибировать средние гены. В свою очередь продуктами средних генов являются два белка gp33 и gp34. Последние вытесняют gp28, образуя комплекс a2ßß’gp33gp34, который осуществляет транскрипцию поздних генов.
Таким образом, фаг SPO1 для транскрипции своих генов использует клеточную РНК-полимеразу, способность которой прочитывать те или иные гены зависит от наличия в ее составе определенных фагоспецифических белков.
У фага Т7 транскрипция генов осуществляется как клеточной, так и фагоспецифической РНК-полимеразами. В геноме фага представлены ранние и поздние гены. После попадания в клетку вначале транскрибируются ранние гены. Транскрипцию осуществляет бактериальная РНК-полимераза. Одним из продуктов ранних генов является фагоспецифическая РНК-полимераза, которая транскрибирует поздние гены. Следует отметить, что промоторы ранних и поздних генов отличаются. Если промоторы ранних генов сходны с таковыми бактериальных, то промоторы поздних генов организованы иначе. Продуктом ранних генов является и еще один регулятор транскрипции. Этот белок инактивирует клеточную РНК-полимеразу. В результате чего прекращается транскрипция ранних генов фага.
По-другому осуществляется регуляция транскрипции генома фага N4. В составе зрелых фаговых частиц находится фагоспецифическая РНК-полимераза. Эта РНК-полимераза обеспечивает транскрипцию ранних генов. Среди продуктов ранних генов имеется вторая РНК-полимераза. Она считывает средние гены. Поздние же гены фага, по-видимому, транскрибируются РНК-полимеразой E.coli.
В регуляции экспрессии генов фагов значительная роль принадлежит антитерминации. В связи с этим рассмотрим регуляцию транскрипции некоторых генов фага λ. Фаг λ принадлежит к числу умеренных фагов. Он либо вызывает типичную инфекцию, либо встраивает свой Геном в клеточную хромосому. Транскрипцию генов фага осуществляет клеточная РНК- полимераза. На ранней стадии развития, последняя, взаимодействуя с промотором, транскрибирует ген N. Транскрипция на этой стадии заканчивается в области терминатора (рис. 11.12). Продуктом гена N является белок антитерминатор, который способствует РНК-полимеразе прочитывать терминатор и транскрибировать расположенные за ним гены (рис. 11.12). Таким образом, в присутствии белка N происходит образование протяженных молекул иРНК, Трансляция которых обеспечивает синтез дополнительных белков.
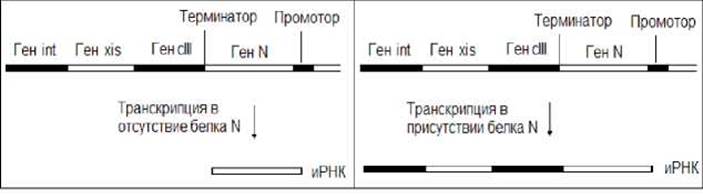
Рис. 11.12. Регуляция экспрессии генов у фага λ посредством антитерминации
Еще один способ регуляции экспрессии генов существует у фага λ. Ген интегразы, белка ответственного за интеграцию генома фага в клеточный геном, может быть транскрибирован с двух различных промоторов. Транскрипт, считанный с одного из промоторов, имеет комплементарные последовательности, благодаря которым формируется двухцепочечная внутримолекулярная структура, которая разрушается РНКазой III. В результате Цистрон, кодирующий интегразу, деградирует и белок не синтезируется (рис. 11.13). Транскрипт же, считанный с другого промотора, стабилен и транслируется с образованием интегразы.

Рис. 11.13. Двухцепочечная структура иРНК разрушается РНКазой III
Транскрипция геномов ДНК-содержащих вирусов эукариот
Эффективность транскрипции генов ДНК-содержащих вирусов эукариот зависит от «силы» промоторов и терминаторов. Гены вирусов эукариот в отличие от генов фагов имеют мозаичную структуру, их экспрессия также может регулироваться энхансерами. Образование иРНК вирусов эукариот включает кэпирование 5’-конца, полиаденилирование 3’-конца, сплайсинг, Альтернативный сплайсинг. Эти модификации часто осуществляются клеточными ферментами.
Транскрипция геномов ДНК-содержащих вирусов эукариот может осуществляться разными РНК-полимеразами. Так транскрипция генома аденовирусов осуществляется в ядре клеточными ферментами. При этом значительная часть генов транскрибируется РНК-полимеразой II, меньшая часть - РНК-полимеразой III. В тоже время транскрипционная система вируса осповакцины локализована в цитоплазме. Это один из самых крупных вирусов, размер его генома составляет около 200 000 п.н. Транскрипционный аппарат, в том числе и вирусспецифическая РНК-полимераза, закодированы в геноме этого вируса. Более того, он находится в сердцевине вирусной частицы. У вируса различают ранние и поздние гены. Промоторы поздних генов отличаются от промоторов ранних генов. Промоторы ранних генов распознает немодифицированная вирусспецифическая РНК-полимераза. Поздние же гены считываются модифицированной РНК-полимеразой. Интересно, что модифицированный фермент включает субъединицу клеточной РНК- полимеразы II. Посттранскрипционная модификация первичных транскриптов осуществляется ферментами, закодированными в вирусном геноме.
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.