Біогеографія - Регіональний аспект - Марисова І.В. - 2005
Глава 2. Характеристика основних кліматично обумовлених груп наземних екосистем та їх біоценозів (біомів)
ЕКВАТОРІАЛЬНО-ТРОПІЧНИЙ ПОЯС
2.1. Вічнозелені екваторіальні та тропічні дощові ліси (гілея)
2.1.3. Рослинність
Вперше ці ліси описав О. Гумбольдт під час своєї подорожі в Південну Америку і назвав їх гілеєю (від грец. hyle — ліс). Рослинність характеризується такими рисами:
1. Різноманіття і специфічність життєвих форм рослин. Тут є дерева, ліани, епіфіти, паразити та ін. Про співвідношення цих форм порівняно з листопадними лісами помірної зони дає уявлення таблиця, складена за класифікацією Раункієра.
Таблиця 2.1. Біологічний спектр лісів різних кліматичних зон (у %)
Типи лісів |
Епіфіти |
Фанерофіти |
Хамефіти |
Гемікриптофіти |
Геофіти |
Терофіти |
Дощові ліси |
22 |
66 |
12 |
0 |
0 |
0 |
Листопадні ліси помірної зони |
0 |
27 |
6 |
39 |
23 |
5 |
2. Надзвичайне багатство і видове різноманітніття. Загальна кількість видів рослин у дощових лісах величезна: на Амазонці - приблизно 40 000 видів; на Зовдських островах - 35 000 видів. Для порівняння скажемо, що в усій північно-східній Євразії налічується трохи більше 17 000 видів.
Дуже багато ендеміків, переважно вигадливих форм і яскраво забарвлених.
3. Полідомінантність. У фітоценозах завжди присутня велика кількість видів. Іноді на 1 га лісу легше знайти 100 різних видів, ніж 100 особин певного виду.
4. Абсолютне флористичне панування деревних рослин. Підраховано, що на 1 га в середньому зустрічається понад 100 видів дерев. Водночас кількість трав не перевищує 20 видів на материках, на островах — 1—3 видів. Таке співвідношення прямо обернене до існуючого в лісах помірної зони. Цікаво відзначити, що родини багатьох рослин, які в помірній зоні є трав'янистими, тут представлені деревами (наприклад, фіалкові, вербенові тощо).
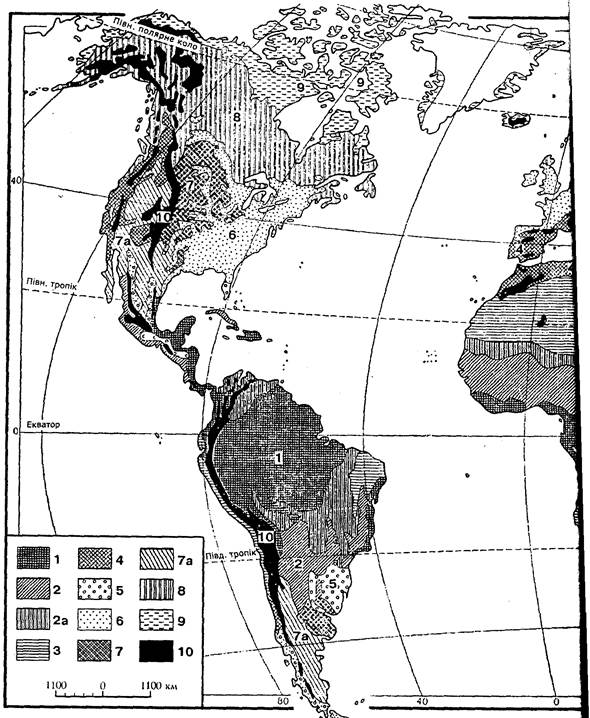
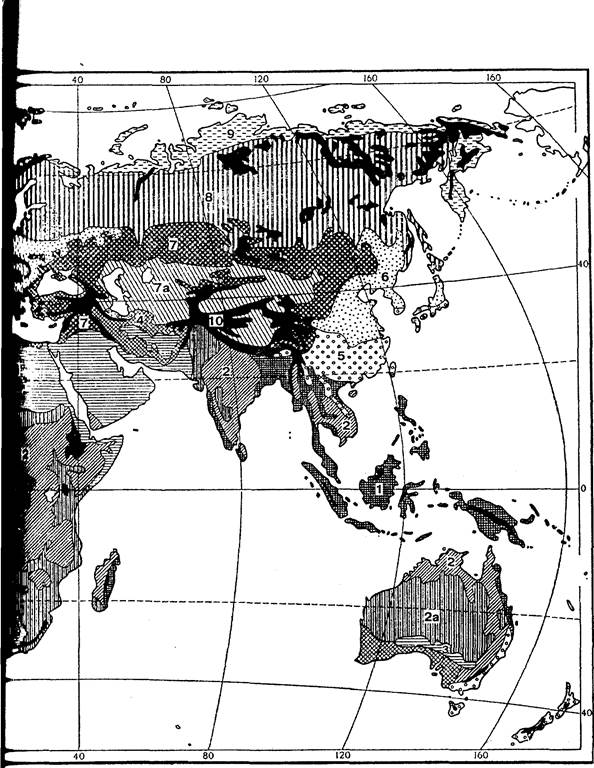
Рис. 1. Схема основних біомів суші (за Г. Вальтером, 1974)
1 - вічнозелені тропічні дощові ліси; 2 - тропічні напіввічнозелені і листопадні ліси; 2а - сухі ліси, рідколісся і савани; 3 - тропічні і субтропічні пустелі і напівпустелі; 4 - шорстколисті середземноморські формації; 5 - вічнозелені листяні ліси помірно теплих областей або постійно вологих прохолодних районів; 6 - літньозелені широколисті (неморальні) ліси; 7 - семиаридні трав’янисті угруповання помірних широт; 7а - пустелі і напівпустелі помірних широт; 8 - хвойні і листяно-хвойні (бореальні) ліси; 9 - тундри і приполярні пустоші; 10 - високогір'я.
5. Невелика площа поширення більшості видів. Здебільшого площа поширення видів обчислюється лише сотнями, а то й десятками квадратних кілометрів. У зв'язку з цим ареали таких видів не виходять за межі тропічної зони. Звідси випливає дуже важливий висновок: знищення рослинності на такій площі призводить до загибелі не тільки окремих популяцій, а й цілих видів, тобто безповоротно скорочується генофонд біосфери. Тому зрозуміло, наскільки вразливий даний біом.
6. Складна вертикальна структура - багатоярусність. Найчастіше в цих лісах розрізняють 4—5 основних ярусів (рис. 2):
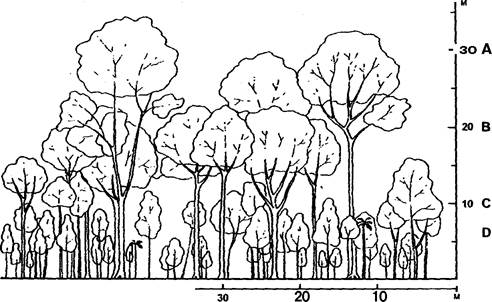
Рис. 2. Вертикальна структура вологого тропічного лісу (за П. Річардсом, 1961)
А - верхній ярус, складається з найбільш високих дерев (до 70-80 м і вище). Вони не утворюють суцільного покриття, і"х крони не стикаються; вони поодиноко або невеликими групами підносяться над суцільним зеленим килимом наступного ярусу. Ці дерева називаються емерджентами; В - другий, або середній деревний ярус, який складається з дерев висотою 20-25 м, іноді до 35 м; крони цих дерев, змикаючись, утворюють суцільний полог; С - нижній деревний ярус, складається з дерев заввишки 12-15 м; Д — ярус молодої порослі дерев і високорослих трав; Е - надґрунтовий ярус трав і сіянців дерев, а також грибів, лишайників тощо.
7. Відсутність чагарникового ярусу.
8. Спільні риси дерев, які складають певний ярус, незалежно від їх систематичної належності: форма крони, характер галуження, розміри, колір, діаметр стовбура, форма і характер
листя тощо. Так, наприклад, всі дерева ярусу А мають парасолькоподібну крону; у ярусі С крона вузька, пірамідальна; у ярусі В крона має проміжну форму.
До того ж у багатьох дерев форма крони різна на різних стадіях онтогенезу. У міру зростання такі дерева поступово досягають вищих ярусів, тобто перебувають у різних екологічних умовах, тому їх крона відповідно змінюється.
Листя дерев одного ярусу також має спільні риси, незалежно від систематичної належності цих дерев. Навіть в одного і того ж дерева листя може бути різним залежно від того, у якому ярусі в даний момент розміщується та чи інша частина крони. Таким чином, листя одного й того ж виду може бути різним, а листя різних видів - однаковим.
9. Наявність «крапельниці» (витягнутого носика на кінці листа в багатьох рослин), що полегшує стікання води з листа.
10. Специфічність молодих листків. Вони дуже різноманітні за забарвленням і різко виділяються на загальному зеленому фоні. Серед них бувають червоні, рожеві, жовті, майже білі, блакитні, яскраво-малинові тощо. До того ж часто таке молоде листя поникле, м'яке, непружне, причиною чого є недорозвинення механічної тканини.
11. Специфічний вигляд великих дерев. Стовбури дерев стрункі, вкриті тонкою, часто зеленою корою, без бічних гілок аж до самої крони.
Від основи стовбура на висоті 8—9 м відходять в усі боки плоскі, так звані дошкоподібні, корені. Вони мають дуже велике значення в житті дерев, виконуючи функцію підпорок, контрфорсів. Річ у тім, що нормальне коріння в таких дерев, у зв'язку з бідністю ґрунтів на мінеральні й органічні сполуки, не заходить у ґрунт глибше ніж на 0,5 м і без бічних коренів-підпорок високі дерева не могли б устояти, а тим більше протистояти частим тропічним ураганам.
Взагалі коренева система дерев дощових лісів розгалужується біля самої поверхні землі, пронизуючи лісову підстилку з опалого листя, відмерлих частин рослин тощо. На цих коренях часто розвивається мікориза (від грец. mykes — гриб ігЬіга — корінь).
12. Наявність кауліфлорїі і раміфлорїі. Це такі явища, коли утворення квітів і плодів іде просто на стовбурах або товстих гілках дерев (прикладом може бути какао). Такі дерева належать переважно до ярусу С або Д. Кауліфлорія полегшує запилення квіток мурашками та іншими комахами, що повзають по стовбурах.
13. Багатство і різноманіття між'ярусної рослинності. Сюди належать ліани та епіфіти.
Ліани - численна життєва форма різних тропічних рослин. Переважна кількість видів ліан є деревними рослинами. Деякі з них (наприклад, фікуси) належать до групи ліан-душителів, які починають свій розвиток як епіфіти, а потім утворюють значну кількість повітряних коренів. Деякі з них обплутують стовбури хазяїна, дерев'яніють і згодом душать його: дерево-хазяїн засихає, а ліана продовжує стояти «на власних ногах». Ліани зустрічаються серед усіх родин тропічних рослин.
Епіфіти - особлива життєва форма рослин, пов’язана з боротьбою між'ярусних рослин за світло і живлення. Особливо багато епіфітів серед папоротей, орхідей, бромелій, плаунів. Серед епіфітів розрізняють:
- епіфіти з цистернами (наприклад, родина бромелієвих), у розетці листків яких збирається і накопичується дощова вода, утворюючи «міні-озерця», де мешкають різні тваринки, навіть рибки;
- епіфіти гніздові та епіфіти-бра (наприклад, оленячий ріг), між корінцями яких накопичуються часточки ґрунту, органічні рештки, що затягуються туди комахами, які повзають по стовбурах дерев.
14. Наявність так званого ефекта Соссюра. Спостерігається в епіфітних кактусів, орхідей і бромелій та полягає в тому, що вдень продихи на листках цих рослин закриті, унаслідок чого транспірація зменшується, що унеможливлює фотосинтез. Продихи відкриваються вночі, коли температура повітря зменшується і рослини накопичують СО2, зв'язуючи його в клітинному соку у вигляді органічних кислот. Наступного дня вуглекислота використовується у фотосинтезі, який здійснюється ніби «за закритими дверима».
15. Відсутність регулярної періодичності в розвитку дерев. Окремі екземпляри дерев одного й того ж виду і навіть окремі гілки того самого дерева цвітуть і плодоносять у різні періоди року, маючи свій власний ритм. Часто можна бачити на одному дереві і квіти, і плоди, і молоде листя.
Те саме стосується особливостей скидання листя окремими деревами (переважно з верхнього ярусу). Через це гілея виправдовує свою назву «вічнозелені ліси», бо ліс ніколи не буває весь голим, як у помірних широтах.
16. Парадокс пишного розвитку дощових лісів на дуже бідних ґрунтах. Цей парадокс обумовлений певними особливостями кругообігу поживних речовин у гілеї. Особливе значення при цьому має своєрідний « ярус живлення », який являє собою прошарок підстилки разом із кореневою системою (підземною і надземною) деревної рослинності.
В екваторіальному кліматі при відмиранні рослин відбувається дуже інтенсивний розклад органіки, елементи мінерального живлення миттєво засвоюються сильно розгалуженим корінням, знову повертаючись у біомасу. Так, понад 90% кальцію, калію, магнію надходить у рослини з біомаси, фактично минаючи ґрунт. До 80% поживних речовин надходить безпосередньо в рослини завдяки мікоризі.
Крім того, важливу роль у кругообігу поживних речовин відіграють своєрідні «мініценози» епіфітів, де постійно накопичується органіка. Вони являють собою проміжні підстилки, які знаходяться на шляху дощової води. Вона проходить крізь них, ніби через кілька фільтрів, від пологу верхнього ярусу дерев до приземного «ярусу живлення», поступово все більше насичуючись поживними елементами.
Отже, умовний «ярус живлення» і фільтрація атмосферної вологи крізь багатоярусний дощовий ліс - найважливіші напрямки надходження поживних речовин у рослини.
У результаті дії цього складного і злагодженого механізму живлення в дощових лісах втрати поживних речовин у ході їх кругообігу є мінімальними. Завдяки цьому механізму рослинність гілеї змогла в умовах стабільності природної обстановки протягом дуже тривалого часу забезпечувати себе необхідним мінеральним живленням і формувати величезну біомасу навіть при незначному вмісті мінеральних елементів у ґрунтах.
Загальна фітомаса первинних дощових лісів досягає в середньому 600-700 т/га, а в деяких регіонах (наприклад, в Амазонській сельві) - 1500 т/га. Річна продуктивність дорівнює 30-50 (нерідко 70) тонн на гектар.
Таким чином, біом гілея - найбагатший і найпродуктивніший на Землі, який сформувався за мільйони років розвитку, унаслідок чого склались екосистеми з найбільш складною структурою і численними міжвидовими зв'язками. Тому випадання з них навіть окремих ланок завжди може стати початком необоротної деградації всієї екосистеми.
Отже, аналіз структурно-функціональних особливостей гілеї свідчить про виключну вразливість цього угруповання і надзвичайну складність його відновлення. Вторинні ліси ніколи не досягають (і не можуть досягнути) багатства первинної гілеї. Це необхідно враховувати при розробці будь-яких проектів природокористування в постійно вологих тропіках. На сучасному етапі гілею слід розглядати як непоновні ресурси біосфери і з урахуванням цього факту планувати її господарське освоєння.
Не можна забувати, що величезні масиви зелених лісів тропіків - найпотужніший генератор кисню на Землі, це «легені» Землі, від стану яких залежить здоров’я людей і саме життя на планеті.
Тепер має стати зрозумілим, чому така значна увага міжнародної спільноти приділяється проблемі збереження вологих дощових вічнозелених лісів тропіків. А те, що найперший проект програми МАБ присвячений цим лісам, підкреслює пріоритетність проблеми.