БОТАНІКА З ОСНОВАМИ ГІДРОБОТАНІКИ (ВОДНІ РОСЛИНИ УКРАЇНИ) - Б.Є. Якубенко - 2011
VІІІ. ОСНОВИ МОРФОЛОГІЇ І АНАТОМІЇ РОСЛИН
Морфологія і анатомія листка
Анатомічна будова листка
Внутрішня будова листка тісно пов'язана з функцією та умовами місцезростання. Типовий листок двосім'ядольних рослин має дорзовентральну будову (верхній і нижній бік вирізняються між собою). Деякі рослини мають ізолатеральну будову листка.
Ззовні плоский листок укритий епідермісом - первинною покривною тканиною, що регулює транспірацію та газообмін, захищає листок від висихання та пошкоджень. Верхній епідерміс одношаровий, зрідка дво- і тришаровий (фікус). Клітини епідермісу паренхімні, щільно зімкнені між собою, з хвилястими контурами, або мають виступи, що сприяють щільному з'єднанню їх між собою. Клітини з густим протопластом, як правило, без хлоропластів, у них часто трапляються лейкопласти. Оболонка клітин епідермісу потовщена не рівномірно. Найпотовщеніші зовнішні і радіальні стінки. Клітини можуть містити пігменти групи антоціанів, які забарвлюють листкову пластинку в різні кольори. В епідермісі утворюється значна кількість продихів (у замикаючих клітинах яких є хлоропласти), які забезпечують газообмін і транспірацію. Переважно вони утворюються на нижній стороні листка, а у водних рослин навпаки, на верхній.
Поверхня епідермісу часто вкрита різноманітними утвореннями. шипами, волосками. Своєю формою, будовою та розмірами вони становлять характерну ознаку для певних систематичних груп рослин. Волоски мають важливе фізіологічне значення, захищаючи покривну тканину листка. У всіх рослин епідерміс зверху вкритий шаром кутикули.
Нижній епідерміс має слабко розвинену кутикулу, продихів багато, вкритий волосками. Клітини нижнього епідермісу, зазвичай, дрібніші та зі звивистиміши клітинними оболонками, у порівнянні з клітинами верхнього епідермісу.
Епідерміс виконує захисну функцію, а наявність продихів сприяє обміну речовин з навколишнім середовищем. Про його інтенсивність можна судити з того, що на 1 мм1 поверхні епідермісу пшениці знаходиться 50-70 продихів, яблуні - 250, картоплі - 263, лимона - 623, традесканції - 7 (на верхньому боці листка). У водних рослин листки яких плавають на поверхні води, продихи розміщуються тільки з верхнього боку, з нижнього, зануреного у воду, вони відсутні. Підводні листки продихів не мають.
Основну тканину листка називають мезофілом Він знаходиться між верхнім і нижнім епідермісом та складається з паренхімних клітин з тонкими й ніжними оболонками, з протопластом і зеленими пластидами. Ці клітини становлять хлорофілоносну паренхіму або хлоренхіму. У клітинах мезофілу вирізняють палісадну (стовпчасту) та губчасту паренхіму.
Клітини палісадної паренхіми видовжені, циліндричні, щільно прилягають одна до одної, мають багато хлоропластів. Вони розміщені одним або кількома правильними компактними рядами. Палісадна тканина становить основну ассиміляційну тканину, чим пояснюється її розміщення на верхній стороні листка.
Клітини губчастої паренхіми мають неправильну форму і пухке розміщення. У товщі мезофілу губчаста паренхіма переважає, але хлоропластів у ній менше у порівнянні з палісадною. Більша частина поверхні клітин губчастої паренхіми межує з міжклітинниками. Будова типової губчастої паренхіми зумовлена її основною функцією - транспірацією. Кількість шарів варіює від 2-7 і залежить від умов середовища. Форма клітин губчастої паренхіми може бути округла, звивиста, лопатева або зірчаста. Великі міжклітинники пронизують цю тканину в різних напрямках. Верхні клітини губчастої паренхіми утворюють спеціальний ряд збиральних клітин, які мають форму лійок, і прилягяють одночасно до кількох клітин палісадної паренхіми. У них накопичуються продукти фотосинтезу, що утворились у клітинах стовпчастої паренхіми.
Мезофіл листка пронизує система жилок. Провідна тканина пластинки та черешка листка становить одне ціле з провідною системою стебла. Усі частини цієї системи утворюються з прокамбію. За утворення провідної системи листка розвиваються такі самі типи клітин. Центральна жилка складається із ксилеми, орієнтованої доверху, і флоеми, поверненої донизу. Подібне розташування ксилеми і флоеми обумовлено виходом цих тканин із стебла і поворотом їх на 90 градусів у листковій пластинці. Ксилема, яка в стеблі займала центральне положення, виявилась назовні, а флоема, розміщена в стеблі з периферії, змістилася донизу. Ксилема складається із судин або трахеїд, ксилемної паренхіми і радіальних променів. За ксилемою у двосім’ядольних рослин міститься камбій, який забезпечує потовщення жилок. Нижче від камбію знаходиться флоема. Вона складається із ситоподібних трубок, клітин-супутниць і флоемної паренхіми. Оточує провідний пучок склеренхімна Піхва. Над провідним пучком і під ним розвинена пластинкова або куткова коленхіма.
Бічні провідні пучки (жилки) та їхні відгалуження мають спрощену будову, часто вони позбавлені механічної Тканини і флоеми. Країв листкової пластинки досягають ксилемні провідні пучки або тільки їхні елементи - судини чи трахеїди.
Таким чином у листках формуються провідні пучки колатерального типу, у більшості рослин закриті.
Класичним об'єктом для вивчення анатомічної будови листків є листок камелії японської (Camellia japonica L.) (рис 77). Це типовий листок, характерний для представників класу двосім'ядольні. Зверху листок вкритий епідермісом. Клітини епідермісу великі, плоскі майже прозорі. На нижній стороні листка добре видно продихи, під якими знаходиться повітряна порожнина. Епідерміс вкритий плівкою кутикули. Під епідермісом верхньої сторони листка знаходиться асиміляційна тканина стовпчаста паренхіма. Це два ряди видовжених клітин, які щільно прилягають не утворюючи міжклітинників. Клітини заповнені хлоропластами, що зосереджені в постійному шарі цитоплазми. Під клітинами палісадної паренхіми знаходяться лійкоподібні збиральні клітини, через які накопичені продукти фотосинтезу передаються в провідну систему листка. Нижня частина заповнена губчастою паренхімою. Її клітини неправильної форми, з великими міжклітинниками. У мезофілі листка камелії трапляються значних розмірів друзи і кристали щавлево-кислого кальцію.
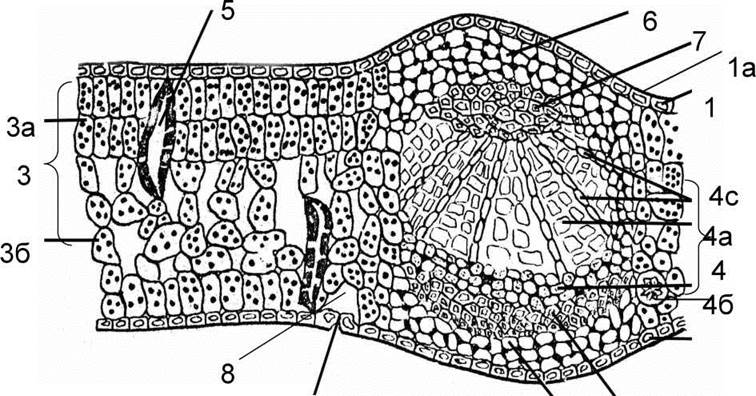
Рис. 77. Поперечний зріз листка камелії японської:
1 - верхній епідерміс, 1а - кутикула; 2 - продих; 3 - мезофіл листка;
За - стовпчаста паренхіма, 36 - губчаста паренхіма; 4 - провідний пучок:
4а - ксилема, 46 - флоема; 4с - радіальний промінь; 5 - опорні клітини;
6 - клітини коленхіми; 7 - клітини склеренхімної обкладки ; 8 - повітряна порожнина.
Провідний пучок захищають кілька шарів пластинково-куткової коленхіми. Вона утворена живими паренхімними клітинами, що мають кутові потовщення і потовщення тангентальних стінок. Коленхіма прилягає до центрального провідного пучка під верхнім і нижнім епідермісом.
Будова листка латаття білого (Nymphaea alba L.). Це водна рослина з великими листками, що плавають на поверхні води. Будова листка дорзовентральна, але в ньому спостерігаються специфічні пристосувальні структури.
Палісадна паренхіма дрібноклітинна і багатошарова. Її клітини відносно щільно прилягають одна до одної, але є вузькі міжклітинники, які розташовані в напрямку довжини клітин палісадної паренхіми, Остання містить велику кількість хлоропластів і забезпечує Фотосинтез. Губчаста паренхіма має великі клітини в якій розміщені великі повітряні порожнини, хлоропластів не значна кількість. На верхньому епідермісі залягає товстий шар кутикули з продихами. Під замикаючими клітинами продиху знаходяться повітряні камери, що тягнуться вздовж трьох-п’яти шарів палісадних клітин і опираються на верхівку однієї з них. Оболонки клітин верхнього епідермісу звивисті, а нижнього - дещо потовщені, конусоподібні, кутикула відсутня. Між живими клітинами епідермісу є мертві клітини, які підтримують живі, що контактують з ними та допомагають разом з повітряними порожнинами підтримувати м'ясисті важкі листки латаття на поверхні води. Судинно-волокнисті пучки розвинені слабко, елементи флоеми і ксилеми дрібні й нечисленні, механічні волокна майже відсутні. Навколо них розвинена пучкова обкладка. У мезофілі листка можна бачити систему гіллястих поодиноких склереїд - опорних клітин, із потовщеними і кутинізованими оболонками.
Анатомічна будова підводного і плаваючого листків глечиків жовтих (Nuphar luteum (L.) Smith.). Листки цієї рослини вирізняються своєю анатомічною будовою. Так, у підводного листка відсутня палісадна паренхіма, мезофіл однорідний, утворений губчастою паренхімою з великими повітряними порожнинами, продихи відсутні. Плаваючий листок дорзовентральний. Палісадна паренхіма якого багатошарова дрібноклітинна, зі значною кількістю хлоропластів. Між паренхіматичними клітинами розміщені великі товстостінні опорні клітини. Губчаста паренхіма має також хлоропласти та великі повітряні порожнини. Продихи розташовані у верхньому епідермісі. Під продиховою щілиною в глибину мезофілу проходять вузькі продихові повітряні камери аж до губчастої паренхіми. Провідні пучки складаються з дрібних елементів ксилеми і флоеми, вони слабко розвинені.
АНАТОМІЧНА БУДОВА ЛИСТКІВ односім’ядольних рослин надзвичайно різноманітна, що пов'язано з пристосуванням до різних умов існування. Листки, багатьох односім’ядольних рослин освітлюються сонцем рівномірно з обох сторін, що зумовлює їхню ізолатеральну будову. Листкові пластинки таких листків як правило вузькі, з паралельним жилкуванням. У листку нижня і верхня сторона подібні і мало вирізняються між собою. Верхній і нижній епідерміс однакові за будовою та кількістю продихів. Мезофіл листка не диференційований на стовпачасту і губчату паренхіму, або ж вона дуже слабко виражена. Якщо стовпчаста паренхіма наявна, то вона виражена слабко і представлена одним рядом витягнутих клітин.
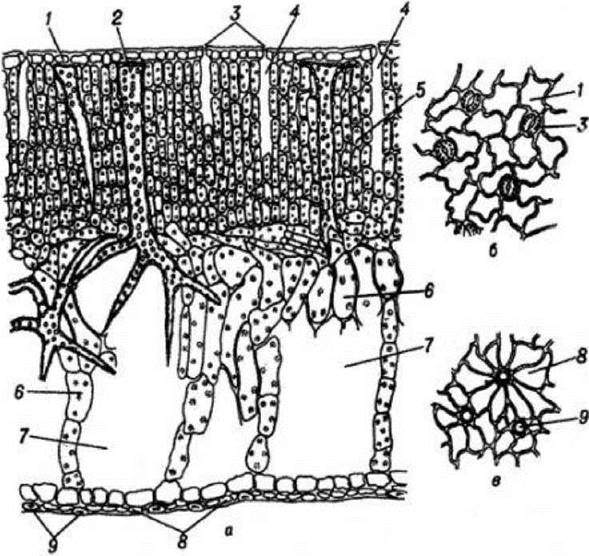
Рис. 78. Поперечний розріз листка латаття білого а — частина листкової пластинки, б - верхній епідерміс з поверхні, в - нижній епідерміс з поверхні, 1 - верхній епідерміс, 2 - опорні гіллясті склереїди,
3 - продихи, 4 - повітряні порожнини, 7 - повітряні порожнини, 8 - нижній епідерміс. 9 - окорковілі клітини нижнього епідермісу.
АНАТОМІЧНА БУДОВА ЛИСТКА півників болотних (Iris pseudacorus L.). З поверхні листок вкритий одношаровим епідермісом (рис 79). Серед клітин нижнього епідермісу трапляються продихи, з двома великими замикаючими клітинами, що містять хлоропласти. Стовпчаста паренхіма складається з одного ряда клітин, знаходиться безпосередньо під епідермісом. У середній частині листка знаходиться оптимально розвинена система міжклітинників, які мають характер повітроносних каналів. На однаковій відстані один від одного з нижнього і верхнього боку листка розташовані в один ряд закриті колатеральні провідні пучки, до складу яких входить флоема і ксилема, оточені склеренхімною обкладкою.
Листки водних рослин мають ряд особливостей анатомічної будови, пов’язаних з умовами існування. Листкова пластинка тонка, прозора, часто розсічена на окремі лінійні частки. Асиміляційна тканина у них розвинена слабко в зв’язку з недостатніми умовами освітлення. Палісадної паренхіми не має, а у губчастої між верхнім і нижнім епідермісом з’являються перемички. Між перемичками формуються великі повітряні порожнини. Епідерміс у вигляді тонесенької плівки, потовщення клітинних стінок епідермісу відсутні. Кутикула надзвичайно тоненька або відсутня взагалі. У епідермісі листкової пластинки, яка повністю занурена у воду продихів не буває, а в деяких водних рослин є особливі утворення, що вирізняються від інших своєю величиною та формою, і більшою проникливістю стінок для води. Вони називаються гідропотами.
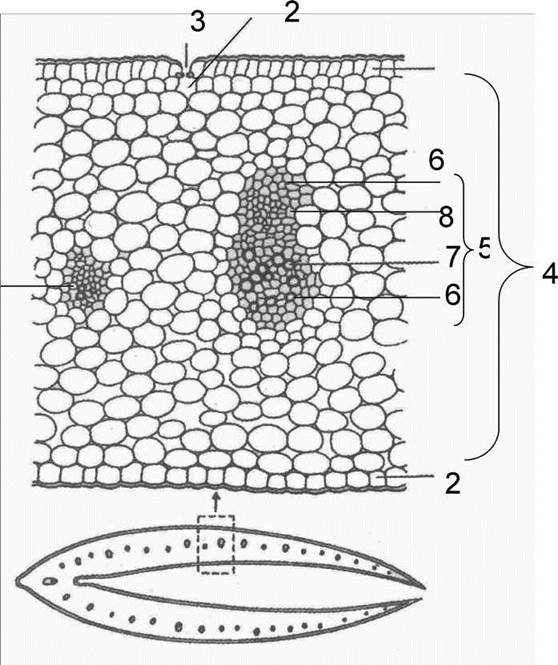
Рис. 79. Поперечний зріз листка півників 1) верхній епідерміс; 2) нижній епідерміс; 3) продих; 4) мезофіллистка; 5) провідний пучок; 6) склеренхіма; 7) ксилема; 8) флоема
У листковій пластинці підводних рослин слабко розвинена ксилема, судинних пучків незначна кількість, судин у них небагато. Часто замість судин у ксилемі наявний вузький міжклітинний хід. Флоема розвинена краще. Система повітряних міжклітинників розвиненою краще, що забезпечує рослину киснем і вуглекислим газом.
У рослин, листки яких плавають на поверхні води, листкова пластинка зазвичай щільна та шкіряста. На верхньому боці якого розвивається значна кількість продихів, під якими знаходиться розвинена повітряна порожнина.
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.