БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
15. РАСТИТЕЛЬНОСТЬ ЗЕМНОГО ШАРА
15.2. Биомы Земли
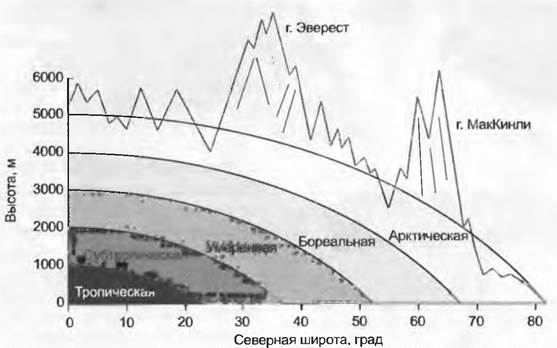
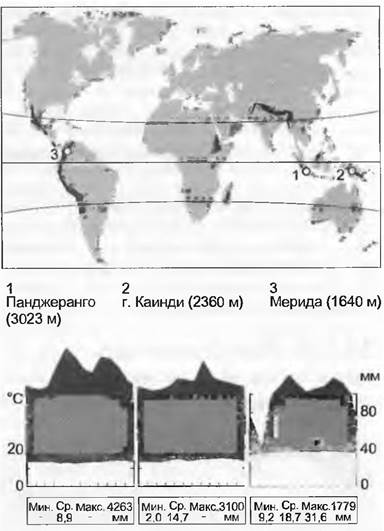
Типичные для климатических зон биомы (зонобиомы) формируются вокруг всего Земного Шара на характерных широтах. В пределах одной широтной зоны различаются гумидные (влажные, атмосферных осадков много), семиаридные (периодически засушливые) и аридные (очень сухие, осадков очень мало) зоны. В пределах всех этих зон, различающихся по температуре и влажности, существуют горные биомы, или оробиомы. Широтной зональности биомов, определяемой температурой и влажностью (но не продолжительностью дня или характером сезонных смен) соответствует высотная поясность в горах (рис. 15.6, 15.7). В горах влажных тропиков можно на небольшом расстоянии пересечь все гумидные температурные зоны Земли. Но из-за различий в сезонных сменах погоды (опасность морозов) растительные зоны соответствуют им очень приблизительно. Это «сжатие» природных зон в тропических горах объясняет их высокое биоразнообразие, поскольку при рассмотрении их в широком плане все высотные пояса объединяются. Отдельные же зоны (высотные пояса) не богаче видами, чем где-либо в другом месте (см. форзац).
Рис. 15.6. Если сравнивать только гумидные регионы, то видно, как с географической широтой и высотой над уровнем моря растительные и климатические зоны сменяются похожим образом. В качестве исходного примера взяты тропические высокогорья, поднимающиеся от пояса влажных тропических лесов до нивального пояса. 1 км по высоте соответствует расстоянию по широте около 2 000 км, 4 км по вертикали до границы лесного пояса с альпийским — расстоянию по широте почти 8000 км от экватора до полярной границы лесов, которая в Европе находится на 68 — 70° с. ш.

Рис. 15.7. Температурные условия (географическая широта) и условия увлажнения формируют растительность Земли: А, В — гумидные и семиаридные тропики; С, D — гумидные и семиаридные субтропики; Е, F — гумидная (океаническая) и семиаридная (континентальная) умеренная зона; G — бореальная и субарктическая зона
В дальнейшем будут рассмотрены 16 биомов; положение их в основном соответствует температурным условиям и количеству осадков на континентах (рис. 15.8).
Хотя горные биомы встречаются во всех климатических зонах, мы рассмотрим только те, которые относятся к умеренной и субтропическо-тропической зонам (в каждом случае горный и альпийский пояса). Описываемые здесь биомы соответствуют следующим обозначениям «зонобиомов» Г. Вальтера: зонобиом (ЗБ) I = 15.2.1, 2, 3; ЗБ II = 4, 5; ЗБ III = 6; ЗБ IV = 7; ЗБ V = 8; ЗБ VI = 9, 10, 11; ЗБ VII = 12, 13; ЗБ VIII = 14; ЗБ IX = 15; представляемые здесь Примеры только ограниченно включают растительность морских побережий (она рассмотрена в разделе 15.2.16), в случае ЗБ I, II, IV, VI.
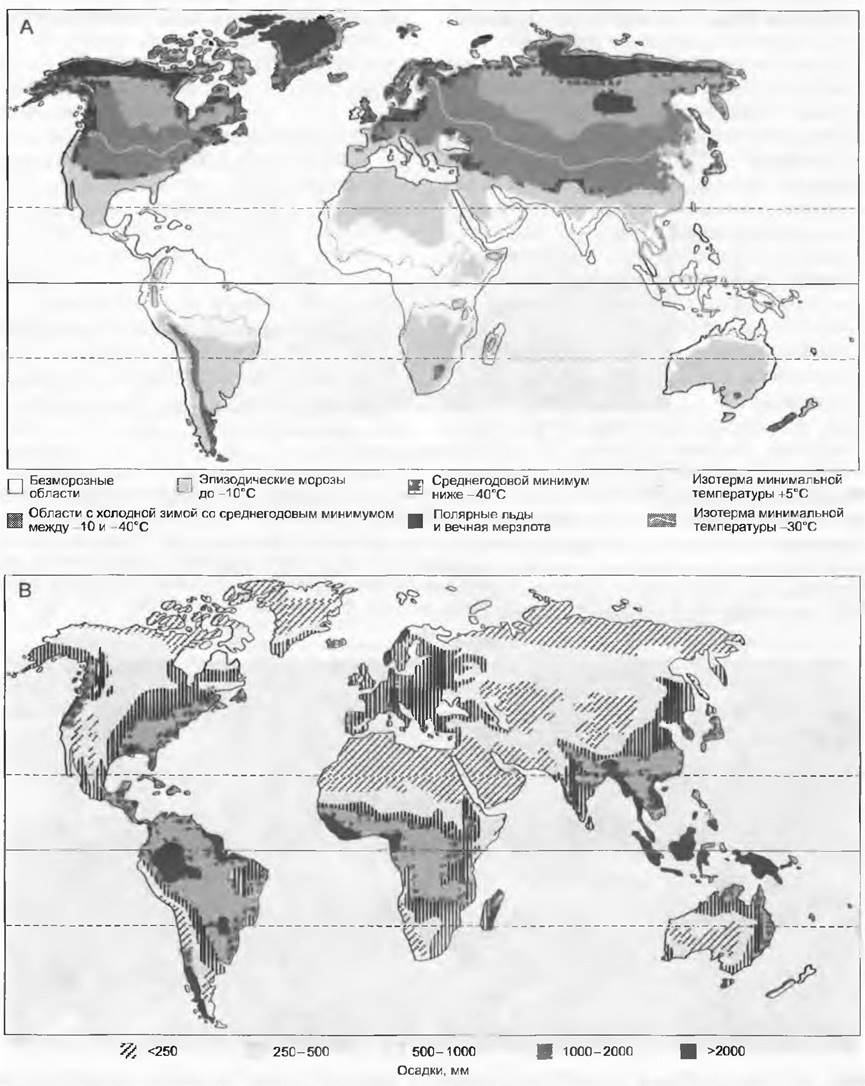
Рис. 15.8. Глобальное распределение двух важнейших метеорологических показателей, определяющих растительный покров Земли: А — максимальное выхолаживание воздуха (морозы); В — годовая сумма осадков. Не принимались во внимание горы и важнейший показатель — сезонность выпадения осадков
15.2.1. Влажно-тропические равнинные леса
Леса сверхгумидных, расположенных близ экватора низменностей (В настоящее время они еще занимают площадь 16 — 17 млн км2, это почти половина всех сомкнутых лесов и около 11 % земной поверхности), не столь однообразны, какими неправильно принято считать дождевые леса. Между ними существуют значительные различия, как флористические, так и климатические, и почвенные. Имеется три крупных тропических гумидных области — север Южной Америки с Амазонией, западная часть Центральной Африки (бассейн Конго и прилегающее побережье) и Юго-Восточная Азия (Южная Индия, Малайзия и Малайский архипелаг, Новая Гвинея, северный выступ Австралии).

Тропические области (расположенные между двумя тропиками) — безморозные области (см. рис. 15.6), представляющие собой климатическую и ботанико-географическую зону, огромную по площади, что следует уже из общей шарообразной формы Земли. Постоянно влажные области занимают только их внутреннюю часть, лежащую примерно в области ± 10° вокруг экватора, или зону ядра. Среднегодовая Температура колеблется здесь в пределах между 24 и 30 °С, годовая сумма осадков составляет от 2 000 до 4000 мм в зависимости от региона. В условиях климата, в котором отсутствуют времена года, даже короткий период без дождя имеет стрессовое значение для эпифитов (суккулентность, САМ-газообмен, устойчивость к высыханию) и дает решающий сигнал для синхронизации цветения у особей одного вида. Циклические климатические феномены, такие, как течение Эль-Ниньо (повторяется раз в 3 — 7, обычно через 5 лет) могут быть причиной длительных засушливых периодов и в Восточной Азии (так же, как и катастрофические дожди на западном побережье Южной Америки).
Тепло и высокая влажность настолько благоприятствуют круговороту веществ в почве, что гумус только едва начинает образовываться и почвы сильно выщелачиваются (образуются оксисоли, или красно-коричневые латеритные почвы, кварцево-песчаные почвы). Запасы минеральных веществ во влажных тропических лесах заключаются в основном в растительной массе (калия — до 90 % от общего запаса в экосистеме), что и вызывает катастрофические последствия выжигания этих лесов. Запасы минеральных веществ, тысячелетиями аккумулировавшиеся в ходе биологического круговорота, минерализуются мгновенно и вымываются дождями. В естественных условиях Минеральные вещества из растительного опада сразу же связываются микроорганизмами и микоризообразующими грибами и потребляются корнями растений (замкнутый круговорот; при этом леса наилучшим образом обеспечиваются питательными веществами, хотя в почвах их почти не содержится). С водным переносом теряется незначительное количество минеральных веществ, но постоянный дальний транспорт осуществляется с переносимой ветром пылью; доказано, что в бассейн Амазонки она попадает даже из Сахары.
Структура древостоя; кроны деревьев находятся на высоте от 30 до 50 м, образуя вместе с эпифитами (виды семейств Вгоmeliaceae, Orchidaceae, Polypodiophyta) сильно разреженный верхний ярус, под которым располагаются субдоминирующие виды деревьев или молодые деревья. Ниже расположен ярус кустарников (например, Piper) и крупных трав(например, Миш, Heliconia) и виды напочвенного яруса. Лианы (вьющиеся, цепляющиеся, лазящие, с корнями-присосками; см. табл. 4.1) пронизывают весь древесный ярус. Некоторые лианы («душители») разрастаются на деревьях после достижения яруса крон настолько сильно, что, убивая их, становятся самостоятельными растениями (это типично для рода Ficus). На листьях поселяются эпифиллы (Водоросли, мхи, Лишайники). Тропические девственные леса представляют собой мозаику различающихся по возрасту насаждений (цв. табл. 15.1). Восстановление их в сильной степени зависит от лиан и эпифитов, под бременем которых деревья падают. Свежие окна в древостое занимают быстрорастущие виды (Cecropia, Ochroma — бальзовое дерево, Musanga, Маcaranga). Деревья более поздних сукцессионных стадий имеют такой же возраст, как и в умеренной зоне (150 — 200 лет). Часто отмечается образование корней-подпорок
из основания ствола (досковидные корни, придаточные ходульные корни у пальм). Листья крон верхнего яруса слегка кожистые (и очень долговечные, что является реакцией на конкуренцию за питательные вещества), большей частью эллиптические и цельнокрайние, и их образование в большом количестве часто имеет такой взрывной характер, что они сначала кажутся вялыми и красноватыми, как бы недоразвитыми («трясущаяся листва»), что это застает врасплох травоядных животных. Многие тропические виды уже при низких положительных температурах (<7°С) получают необратимые повреждения («выхолаживание», см. 13.3.1). Решающий же экологический фактор — конкуренция за свет.
Важные пантропические семейства — Аrасеае (например, Monstera), Аrесасеае (пальмы), Araliaceae (Schefflera), Bignoniaceae, Caesal- piniaceae, Lauraceae, Moraceae (Ficus), Piperaceae, Zingiberaceae и др. Для палеотропической области типичны Dipterocarpaceae (диптерокарповые, имеющие плоды с двумя крыльями) и Pandanaceae (пандановые пальмы); для неотропиков — Bromeliaceae (Tillandsia). На одном гектаре площади можно найти 60 — 100 видов деревьев (рекордное количество в Перу — 300), причем 2/3 из них представлено единичными экземплярами. Такое высокое видовое разнообразие объясняется отсутствием нарушений в прошлом (оледенение, засушливые периоды), отсутствием морозного периода, древностью самих экосистем и изначально их большим, связанным в отдельных частях, ареалом.
15.2.2. Влажно-тропические горные леса
Выше 1 000 — 1 800 м, до высоты 3 000 — 4 000 м над ур. моря расположена зона горных тропических лесов (также их называют горными дождевыми лесами, туманными или облачными лесами) (цв. табл. 15.2). На таких высотах они еще больше обеспечены влагой, поскольку при том же количестве осадков испарение здесь меньше. Почти ежедневно в первой половине дня леса оказываются в зоне конвективной конденсации (туманы — с высоты 1 800 м), в которой остаются до конца второй половины дня, или находятся в зоне постоянно нагоняемых пассатами облаков. В нижней части, близ границы с равнинными лесами, они достигают высоты 45 м, которая на достаточно высоких горах между 3 600 и 4000 м снижается до 3 — 5 м (криволесье).
Среднегодовая температура на высоте 2 000 м составляет около 17 °С (т. е. близка к среднеиюльским значениям для Центральной Европы), на 3 000 м — около 11 °С, а на верхней границе леса только 6 °С. Начиная с 2 500 м возможны слабые заморозки, но это случается очень редко, с 3 000 м — чаще, с 4 000 м они случаются почти каждую ночь (см. 15.2.3). Осадки в нижнем поясе горных лесов превышают 2 000 мм, что при условии очень низкого испарения означает сильное превышение потребности. Это вызывает большой риск развития склоновой эрозии, поэтому для защиты почвы на этих высотах безусловно необходима нормально развитая растительность. Выше зоны конденсации количество осадков падает, но биологически возможного дефицита влаги чаще всего оно не создает.
С увеличением высоты происходит образование мощного слоя грубого гумуса и модергумуса, в котором содержится большое количество питательных веществ, доступных для растений не напрямую. Более прохладные условия и длительное избыточное увлажнение задерживают разложение опада, что. в свою очередь, препятствует развитию напочвенной растительности и возобновлению леса. Из почвы частично выходят на поверхность корни, и находящиеся на них микоризообразующие ГРИБЫ пронизывают свежеопавшую листву (укороченный цикл круговорота питательных веществ).
Сочетание постоянного переувлажнения и полного отсутствия заморозков в нижних поясах горных лесов обусловливают их сходство с равнинными лесами, различия скорее чисто флористические. Для средних высот (1 800 — 2 500 м) свойственна наибольшая плотность пышно развивающихся эпифитов. В верхний пояс эпифитные сосудистые растения почти не идут, замещаясь криптогамными. Разнообразие лиан снижается с высотой над уровнем моря, расчленение на ярусы постепенно исчезает и, наконец, остается только ярус низких деревьев или высоких кустарников, создающих сомкнутый полог крон с небогатым подлеском. Исчезновение горных лесов в пределах своего ареала тесно связано с деятельностью человека. Граница леса опустилась в большинстве случаев на несколько сотен метров. По высокогорным реликтовым лесам (большей частью на устойчивых к пожарам скальных участках, какие-либо другие микроклиматические особенности отсутствуют) можно судить о том, что они способны существовать на высотах около 4000 м. Рассмотренные в разделе 15.2.3 гигантские кроно-розеточные формы указывают на потенциальную возможность существования здесь деревьев.
Тропические горные леса в нижнем поясе очень богаты видами (например, знаменитые леса на уровне облаков на Ранчо Гранде близ Валенсии, Венесуэла, на высоте 1 100 м, см. V.Vareschi). На низких уровнях еще доминируют обычные тропические семейства: пальмы, Moraceae, Rubiaceae и др., на среднем уровне они начинают отступать и на верхнем отсутствуют. С увеличением высоты большую важность приобретают древесные виды следующих семейств: Fagaceae (Castanopsis в Восточной Азии, Quercus в Центральной Америке и ЮгоВосточной Азии), Nothofagaceae на Новой Гвинее, Ericaceae (Erica в Африке, Rhododendron, Vacdnium в Южной и Юго-Восточной Азии), Lauraceae, Myrsinaceae; ближе к границе леса Rosaceae (Polylepis в Южной Америке, Hagenia в Африке) и все Asteraceae. С высотой сильно возрастает роль хвойных и древовидных папоротников (виды Podocarpaceae — Dacrydium, Podocaipus; из древовидных папоротников Суаthea), но они встречаются не везде, нигде не доминируют и большей частью не достигают верхней границы леса. В Коста-Рике количество семейств (и, соответственно, видов) древесных растений падает с высотой от 82 (349 видов) на высоте 2 000 м до 34 (74 вида) на 3 200 м. На самых высоких уровнях (близ верхней границы леса) встречается обычно менее 5 видов, причем бросается в глаза увеличение роли Rosaceae (Южная Америка и Африка) и Ericaceae (Африка и Юго-Восточная Азия). Широко известны в первую очередь островные рощицы из Polylepis на высоте почти 4 000 м в тропических Андах, в экваториальной Африке им соответствуют кустарниковые заросли из Hagenia (оба — представители семейства Rosaceae).
15.2.3. Растительность высокогорий тропиков и субтропиков
Естественная верхняя граница горных лесов и нижняя граница безлесного альпийского (= андского, = афро-альпийского) пояса в экваториальных тропиках лежит между 3 600 и 4000 м над ур. моря, в субтропиках в отдельных районах даже выше. Существенно ниже (до 3 100 м) граница леса — в области высокогорных пустынь, на островах и в таких невысоких горах, где вершины обнажены не по причине неподходящих температурных условий (например, г. Кинабалу). Большая часть высокогорных тропических злаковников («парамос») в настоящее время расположена значительно ниже потенциальной границы леса и возникла в результате антропогенного воздействия (выжигание леса под пастбища).

Климат тропических высокогорий имеет переменный суточный режим с регулярными ночными заморозками в течение всего года и среднемесячными температурами в нижней части от 5 до 7°С. В субтропических высокогорьях имеется смена времен года, выражающаяся в температурном режиме, с морозными малоснежными или бесснежными зимами и несколько более теплым летом, в течение которого также выпадают осадки. Осадков здесь обычно меньше, чем в поясе горных лесов (даже в сверхгумидных районах), обычно менее I 500 мм, в отдельных районах, прежде всего в южных Андах, — менее 500 мм, на таких же высотах в отдельных районах Юго-Восточной Азии, где застаиваются облака — более 3 000 мм (например, г. Вильгельма на Новой Гвинее).
Почвы под сомкнутой растительностью во влажных районах заболоченные, черные. В сухих областях под более редкой растительностью большей частью обычны слабо развитые примитивные почвы (щебнисто-иловато-суглинистые или песчаные). В отличие от предыдущих представлений, произрастающим здесь растениям едва хватает влаги для покрытия физиологического дефицита, только если за год выпадает более 350 мм осадков. Изреживание растительности в таких районах (листовой индекс заметно меньше 1,0), казалось бы, предотвращает возможный в данной ситуации перерасход влаги, при этом остается неясным, каким образом регулируется плотность сообщества. Серьезным препятствием для закрепления молодых растений на обнаженных участках грунта служит регулярно образующийся по ночам игольчатый (стебельковый) лед в верхних сантиметрах почвенного профиля.
Господствующий тип растительности — злаковники с отдельно торчащими разветвленными кустарниками, так называемый «парамос». Во влажных областях экваториальных тропиков доминируют мощные дерновинные злаки (англ. tussock grasslands), в более сухих районах злаки корневищные, формирующие клональные «гирлянды» (в очень сухих, более южных внутренних Андах — почти чистые заросли низких кустарников, «пуна»). Примечательный элемент тропических (но не субтропических) высокогорий — конвергентно сформировавшиеся гигантские розетки, часто эти кроново-розеточные растения (цв. табл. 15.3) выглядят как древовидные формы: в Африке это Dendrosenecio (Asteraceae) и Lobelia (Lobeliaceae), в Андах — Espeletia (Asteraceae) и Puya (Bromeliaceae), на Гавайях — Argyroxiphium (Asteraceae).Такие древовидные виды могут достигать высоты 6 м. Листовые розетки могут закрываться на ночь (защита верхушечной ростовой зоны от радиационного выхолаживания), отмершие основания листьев у молодых экземпляров защищают влагу, содержащуюся в стебле, от ночного замерзания. Но поскольку более старые экземпляры обходятся без такой защиты, она представляется не столь уж существенной. Туника из листьев истолковывается также, как защита от выгорания. Кроновые розетки у Asteraceae обычно плотно беловойлочно опушенные, они имеют много общего с опушенными свечеобразными растениями из родов Lupinus (Анды) и Saussurea (Гималаи). Густое опушение защищает от излучения и смачивания, хотя большое число горных растений тропиков и субтропиков обходятся без такой защиты. От ультрафиолетового излучения внутренние части растений хорошо предохраняют обильно содержащиеся в эпидермисе защитные вещества (например, флавоноиды). Многие тропические альпийские растения защищены от замерзания путем переохлаждения (см. 13.3). Растения-подушки на удивление редки и некоторую роль играют только в отдельных частях южных Анд (например, Azorella, Apiaceae) [и в горах Эфиопии (виды Helichrysum, Asteraceae) — Ред.], что объясняется слабым ветровым воздействием на растения в горах тропиков и субтропиков.
Флористический состав растительности тем более сходен во всех высокогорьях мира, чем выше они расположены. Наиболее существенную роль в сомкнутых сообществах всех высокогорий, включая тропические и субтропические, играют семейства Роасеае и Asteraceae. Всюду встречаются представители родов Festuca и Роа, Сагех, Gentiana / Gentianella, Senecio и некоторые близкородственные роды семейства Ericaceae (например, Vaccinium/ Gaultheria/ Pemettya). Из низких кустарников важную роль играет род Hypericum (Африка, Анды), в то время как в южных Гималаях и в Индонезии сходные экологические ниши заняты видами Rhododendron.
15.2.4. Тропические полувечнозеленые леса
Вдоль краевых частей тропиков выпадение осадков носит типичный сезонный характер, что определяет и сезонный характер растительности (леса, облиственные в период дождей, муссонные леса, сухие леса). Смены дождливого и засушливого периодов приводят к периодическому сбрасыванию листвы. Сезонно-зеленые тропические леса, если бы они не были нарушены, доминировали бы на значительной части планеты (потенциально они занимают 42 % площади всех тропических лесов, или около 7 млн км2). Локальное экранирование лесов горами в тропиках может привести к образованию сухих лесов. Саванны, имеющие сходные климатические ритмы, будут охарактеризованы в разделе 15.2.5.

Климат, для которого характерно периодическое изменение влажности, возникает вследствие периодической смены термических условий по временам года по разные стороны экватора. В течение северного лета экваториальная зона дождей смещается к северу, а южным летом — к югу. Эта астрономическая регуляция усиливается в отдельных регионах связанными с ней атмосферными потоками; влажные воздушные массы, идущие по направлению к полюсу, перемещаются в сторону востока (пассаты). Градиенты температуры и давления между холодным морем и огромными жаркими внутренними районами Азии обусловливают во время северного лета перенос влаги по направлению к суше (муссоны). Время начала (примерно в июне) и интенсивность периода муссонов варьируют, это имеет значительные экологические и хозяйственные последствия. «Зимнее» полугодие бедно осадками, или вообще полностью засушливое. Годовые суммы осадков при равномерном их распределении чаще всего были бы вполне достаточны и для постоянно зеленых лесов. В некоторых регионах годовое количество осадков колеблется вокруг значения 1 500 мм, но при высокой энергии испарения в атмосферу в этих широтах могут существовать только сухие леса. Среднегодовая температура на равнинах соответствует таковой для сверхгумидных тропиков (24 — 30 °С), но с постепенным удалением от экватора строго подчиняется сезонности (сухой период холоднее, влажный — теплее).
Почв формируются под воздействием характерной смены чрезмерной влажности и еще большей сухости, но, за исключением участков с наносным грунтом, они, как и в очень влажных центральных частях тропиков, представлены большей частью сильно выщелоченными оксисолями. С увеличением сухости все более решающее значение приобретает водоудерживающая способность почв. Песчаные и примитивные почвы чрезвычайно усиливают действие засухи (например, каатинга в Венесуэле). Часто имеется плотный подпорный горизонт (железистый, кремнеземный, карбонатный). Поскольку основная часть сообществ теряет листву к началу сухого периода, и при этом к началу следующего периода дождей переработка опала путем обычной деятельности микроорганизмов не может осуществиться, то с возрастанием сухости все более важной в минерализации органики становится роль термитов и пожаров.
Охарактеризовать в едином плане этот биом невозможно, поскольку часто на небольшом пространстве существует вся гамма переходов — от влажного тропического леса до аридных колючих кустарников. Характер конкретного леса определяется длительностью засушливого периода и количеством осадков. С увеличением длительности засухи снижается высота деревьев, исчезают эпифиты и лианы, и в большей степени проявляется дифференциация фенологических ритмов. Опадение листвы происходит в несколько этапов, у некоторых листопадных видов оно происходит всегда, и рано (например, у Bombacaceae), другие виды сбрасывают листья позже, а иногда не теряют их вообще. Во многих муссонных лесах подлесок всегда остается зеленым. Цветение почти всегда приходится на период дождей, но характерные спектры цветения существуют для каждой фазы годичного цикла. Некоторые виды цветут даже в середине засушливого периода. Корни растений уходят на глубину до 30 м и более. Полувечнозеленые тропические леса в настоящее время в результате деятельности человека находятся под еще большей угрозой, чем влажные тропические леса, и площадь их сильно сократилась. Их легкая возгораемость во время сухого периода значительно упрощает корчевку. К тому же большие площади этих потенциально лесных территорий находятся в плотно населенных регионах (например, Индия, окраины тропических областей Африки).
Видовое разнообразие тропических полувечнозеленых лесов чрезвычайно велико, что обусловлено многообразием функционально различных групп видов, иногда даже выше, чем в сверхгумидных областях (ив. табл. 15.4). Спектр ведущих семейств для разных районов меняется. В Южной Америке это виды Bombacaceae с бутылочными стволами (Chorisia), затем Burseraceae, Bignoniaceae (Tabebuja), Anacardiaceae (Гран-Чако — Schinopsis = дерево квебрахо). В Юго-Восточной Азии — муссонные леса с Verbenaceae (Tectona grandis =тиковое дерево), Dipterocarpaceae (Shorea robusta), Combretaceae (Terminaba sp.). В Африке, например, в ксерофильных лесах «миомбо» — на поздних стадиях сукцессии — Caesalpiniaceae (Julbemardia, Brachystegia), на ранних — Terminaba.
15.2.5. Тропические саванны
Саванновая область Земли (около 15 млн км2) по типу сезонной ритмики, обусловленной водным режимом, близка к тропическим полувечнозеленым лесам. Однако под совместным действием климата, почв, пожаров и диких зверей здесь сформировалась специфическая растительность злаковников, перемежающихся редкими лесами, густыми чашами и галерейными лесами вдоль водотоков (цв. табл. 15.5). В Южном полушарии саванны господствуют от тропиков до южной окраины субтропиков, и занимают в Африке наибольшие площади. Аналогичные формы растительности — льяносы Ориноко, серрадос и часть равнин Гран-Чако в Бразилии и бассейне Параны, имеются они и в северной Австралии.

Климат саванн представляет собой еще более сухой вариант климата полувечнозеленых сезонных лесов (см. 15.2.4), но годовое количество осадков здесь, как правило, меньше 1 500 мм, иногда менее 1 000 мм, и здесь более выражено его варьирование по годам, что невозможно отразить на климадиаграмме. При сумме осадков менее 500 мм саванны сменяются полупустынями. В Африке, благодаря наличию высоко приподнятой древней гондванской платформы, на южных окраинах области возможны заморозки. Ночные температуры в течение зимнего полугодия там постоянно держатся ниже 10°С.
Характер почвенного покрова и связанной с ним мозаики растительности саванн в значительной степени обусловлен микрорельефом В Южной Африке постоянные чередования приподнятых плоских холмов и депрессий впадин (часто отличающихся по высоте на 1 или несколько метров) создают мозаику (1) сухих, бедных питательными веществами, сильно выветренных и кислых почв на повышениях, (2) более влажных, суглинистых, богатых почв лощин, со значениями pH до 9 и более, и (3) полностью выщелоченных песков на склонах, на местах выхода инфильтрационных вод по верхнему краю глинистых впадин (англ. seapline) или латеритного водоупорного горизонта. В бразильских серрадос, венесуэльских и колумбийских льяносах и в саваннах северной Австралии такая же мелкоконтурная мозаика почв, различающихся по влажности и степени коркообразования («arecife» в Южной Америке) накладывает отпечаток на их богатство и растительность. Большую роль в круговороте питательных веществ играют термиты и пожары.
Африканские саванны — это разреженные древесные насаждения, которые при отсутствии пожаров и воздействии стад слонов и крупных копытных могли бы за короткое время «зарасти». Локальные нарушения лесов слонами открывают экосистему, создавая возможности для питания животных, обгрызающих низкие ветви (например, антилопы-импалы), и выпаса травоядных (зебры, гну). Это, в свою очередь, препятствует повторному возобновлению леса и в то же время создает условия для возгорания травы, что и создает облик саванн. Чем больше злаков, тем чаще пожары (каждые 2 — 3 года, но часто и ежегодно) и меньше возможностей для роста деревьев. Естественные пожары в саваннах большей частью ежегодно выносят в атмосферу около 1,4 гигатонн (=109 т) углерода, это значительно большее количество СO2, чем выносится от тропических (0,5) и всех прочих (0,2) лесов вместе (в случае с саваннами это часть естественного круговорота углерода, у тропических лесов — чистая потеря, вынос в атмосферные потоки). По причине пожаров в почвах отсутствует подстилка, на ней более интенсивно образуется корка, что препятствует просачиванию дождевой воды и увеличивает поверхностный сток. Величина популяций копытных (зависящая от разнообразия пиши, дождей и крупных хищников, таких, как львы и леопарды) регулирует взаимоотношения леса и злаковников. Пожары были случайно спровоцированы еще первыми гоминидами предположительно более 1 млн лет назад. Неоправданная борьба с ними, происходящая от неправильного понимания природоохранных задач, как и вмешательство в отношения популяций копытных и хищников, может нарушить в этой хрупкой системе баланс между чистыми злаковниками и сомкнутыми сухими лесами.
Если не принимать во внимание флористически богатые галерейные леса, можно выделить три главных компонента африканских саванн: на равнине и в понижениях С4-злаки(например, Pennisetum) и очень колючие виды рода Acacia (Mimosaceae), а на повышениях различные Combretaceae (Combretum sp. с характерными четырехкрылыми плодами). Важные элементы льяносов — Curatella, Byrsonima и др., в районе Чако — Prosopis, Aspidosperma, Schinopsis, пальмы рода Copernicia и др., в северной Австралии — вечнозеленые виды Eucalyptus и неколючие виды Acacia с филлодиями вместо перистых листьев (или безлистные). Для Австралии также типичны такие конвергентные формы деревьев с суккулентными стволами, как Brachychyton (Sterculiaceae), для Африки — Adansonia (баобаб, Bombacaceae), как в Африке, так и на о. Сокотра Dracaena (драконово дерево, Dracaenасеае), в Южной Америке — Chorisia (Bombacaceae, см. 15.2.4).
15.2.6. Растительность жарких пустынь
В обоих полушариях между линиями тропиков и умеренной зоной (или средиземноморской областью зимних дождей) лежат большие засушливые области, называемые пустынями и полупустынями. Своим возникновением они обязаны ориентированному к экватору обратному воздушному течению, которое в процессе передвижения поднимает экваториальные дождевые облака к зениту и обезвоживает воздушные массы (см. рис. 12.7). По сути эти области охватывают пустыни Мексики и Аризоны (Сонора), полупустыни югозападной Бразилии и северо-западной Аргентины, Сахару и Аравийскую пустыню, части северо-западной Индии и Пакистана, Карру в южной Африке и сухие области Центральной Австралии. Особые засушливые зоны, возникшие под влиянием холодных прибрежных течений, имеются на юге Перу — севере Чили (Атакама) и в юго-западной Африке (Намиб).

Эти засушливые субтропические области получают от 0 (Атакама) до примерно 250 мм осадков в год. В отдельных регионах выпадают сезонные частые дожди, как в южной части Сахары в течение северного лета или в северной части зоны пустынь южного полушария, тоже летом, но они редко превышают 100 мм. Существуют климатологические доказательства того, что почти полное разрушение тропических лесов на западе Африки произошло вследствие наступления сухих вихрей из южной Сахары. Пустыня Сонора находится на пересечении областей муссонного (летнего) и средиземноморского (зимнего) типа осадков. Температурный режим здесь имеет выраженную сезонность лето очень жаркое, а зимние температуры низкие, с возможными заморозками.
Почвы очень слаборазвитые или примитивные. Из-за постоянного пересыхания они обогащены в поверхностном слое солями щелочных металлов (или гипсом), что приводит к формированию крайне щелочных (pH>10) почв, особенно в понижениях. По характеру субстрата в Сахаре традиционно различают следующие типы: каменистые пустыни (гаммады), галечниковощебнистые пустыни (реги) и песчаные пустыни (эрги), а также разные формы засолённых пустынь вплоть до совершенно лишенной растительности соляной корки (шотты, засолённые впадины). Усиленная хозяйственная деятельность в прилежащих областях побережья может привести к антропогенному опустыниванию (синдром Сахеля). Существенный фактор — глубина залегания грунтовых вод. Наличие посреди пустыни деревьев указывает на выходы грунтовых вод, причем корни могут доходить до глубины>50 м (например, Prosopis в Неотропической области, Acacia в Африке; см. табл. 13.3 в разделе 13.7.5.1).
В зависимости от увлажнения растительный покров пустынной области изменяется от почти полного отсутствия, это лишайниковые пустыни, или эфемеровой растительности (однолетники, живущие недолго, и появляющиеся только в сезоны с хорошими дождями) до разреженных мимозовых низколесий (максимальная высота их — до 8 м, сложены высокими кустами видов Prosopis или Acacia), а при высокостоящих грунтовых водах — до оазисов с лесами из тамариска (Tamarix) и финиковых пальм (Phoenix). В отношении биомассы в целом важнейшими компонентами являются низкорослые одревесневшие формы (кустарники) с очень глубокой корневой системой (цв. табл. 15.6), причем соответственно уровню грунтовых вод произрастают либо периодически облиственные (многие Mimosaceae), либо длительно зеленые формы — например, виды рода Larrea(креозотовый куст, семейство Zygophyllaceae), распространенные от Северной Америки (L. íridentata), до Южной (L. divaricata). Эти специфические пустынные растения не испытывают неизбежного стресса, их присутствие — следствие дефицита влаги (в другом случае здесь выросли бы другие растения). Сильное изреживание сообществ и приспособленные к особым условиям фенологические ритмы регулируются водным режимом. В активном состоянии эти растения ассимилируют и транспирируют отчасти даже больше, чем такие же в гумидных областях. Суккуленты, чье существование обеспечивается собственными запасами влаги (только у относительно мелко укореняющихся растений), ограничены скорее более влажными районами этой засушливой области. Самое крупное царство суккулентов находится в пустынях Мексики и Аризоны, где осадков очень мало, но они выпадают регулярно (см. выше). Существенную роль играют также терофиты и геофиты, которые зеленеют и цветут только после выпадения дождей, а в песчаных пустынях также вегетативно разрастающиеся клональные злаки (например, Aristida púngeos, злак из Сахары).
Флористически эта пустынная зона скорее бедна. Бросается в глаза повсеместное присутствие Mimosaceae (Acacia, Prosopis, Cercidium), Zygophyllaceae (парнолистниковые — Larrea, Zygophylium), Solanaceae (Lycium) и при наличии засоления Clienopodiaceae (Atripiex, Suaeda). Странный курьез представляет собой древнейшее растение Welwitschiamirabitis, произрастающее в пустыне Намиб (см. рис. 11.211). Среди суккулентов наблюдается необычная конвергенция между стволовыми суккулентами семейства CactaceaeНеотропической и Euphorbiaceae Палеотропической областей, и аналогично — между листовыми суккулентами Agavaceae и Asphodelaceae (Liliales, Aloe sp.). Как Cactaceae(например, Carnegia, Cereus), так и Euphorbiaceae (Euphorbia sp.) представлены одревесневшими растениями до 10 м высотой. В Южной Африке представлено большое разнообразие стволовых суккулентов семейства Asclepiadaceae (Ceropegia, Stapelia и др.) и листовых суккулентов из Aizoaceae (Mesembryanthemum, хрустальная трава, LIthops,живые камни).
15.2.7. Область зимне-зеленых лесов средиземноморского климата
Область, пограничная между умеренным и субтропическим климатом, соответствует климату средиземноморского типа и характеризуется вечнозеленой жестколистной растительностью. Наиболее выражена эта климатическая зона в области Средиземноморья, аналогичные зоны имеются в Калифорнии и Чили, в Капской области и в Австралии.
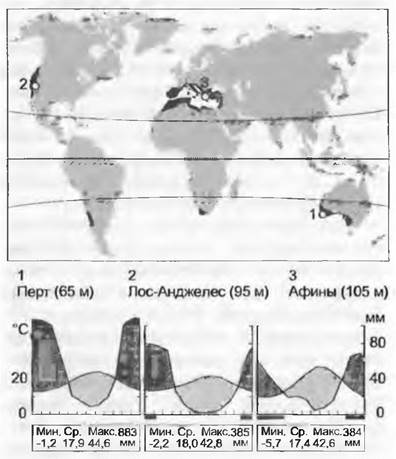
В летнее время эта зона попадает в сухой субтропический пояс, сдвигающийся в направлении полюса, зимой — в зону с умеренным климатом, обусловленным западными ветрами. Годовая сумма осадков лежит между 400 и 1 100 мм (обычно 500 — 800 мм), причем в Средиземноморье большая часть их выпадает в период с ноября по февраль. Зимой на уровне моря возможны заморозки до -6 °С (на севере до -14 °С, гибель олив в Тоскане). Летние температуры регулярно достигают 35 °С и более. Западное Средиземноморье еще испытывает влияние Атлантики, а восточное (Греция, Турция, страны Леванта) более континентальное (осадков здесь меньше, а летние температуры более высокие).
Системы ветров определяют климат регионов — это. например, холодные, часто приносящие летние дожди резкие порывистые восточные ветры в северной Адриатике (бора), а также сухие и горячие очень порывистые северные ветры в восточном Средиземноморье (этезии), дующие в середине лета, которые создают встречные потоки переднеазиатским муссонам (средиземноморский климат часто называют этезийным климатом).
На аллювиальных отложениях низменностей часты погребенные химически выветрелые почвы. Они относятся к группе буроземов (камбисолей) или парабуроземов (лювисолей, образованных глинистыми отложениями, см. 12.5.2.3), на известняках они окрашены в красный цвет (так называемая терра росса). Также часто встречаются слаборазвитые гумусные почвы, лежащие непосредственно на скальных подстилающих породах (рендзины и ранкеры). Выживанию растительности в течение засушливого периода способствуют глубоко проникающие трещины в горных породах с мелкоземистым материалом (корни достигают>20 м в глубину).
Для этого типа растительности свойственно наличие плотных, длительно живущих листьев («жестколистность», склерофиллия). Без каких-либо доказанных причинных связей их часто называют ксероморфными (т. е. обусловленными сухостью), хотя склероморфные листья имеются у растений в каждой из климатических зон, даже в Арктике. Скорее, склерофиллия связана с долговечностью листьев, сильным прессом со стороны травоядных и снабжением их питательными веществами (не прямым образом также и с влажностью почвы), см. 13.6.3. Жестколистность западноавстралийского скраба объясняется также дефицитом питательных веществ (особенно фосфора). Наличие летнее-зеленых видов (в Средиземноморье Fraxinus ornus, Paliurus spinacristi и др.) подтверждает, что этот тип может функционировать несмотря на сухость. В средиземноморских областях северного полушария климаксовая растительность была бы представлена вечнозелеными дубовыми лесами (Quercus agrifolia в Калифорнии, Q. ilex в Европе). Из-за высокой (40 — 100-летние циклы) частоты пожаров (частично вызванной антропогенными факторами) условия здесь очень благоприятствовали развитию пирофильных видов сосен (Pinus). При пожарах, повторяющихся чаше, чем раз в 40 лет, развиваются сообщества жестколистных кустарников (маквис в Средиземноморье, чапараль в Калифорнии, матораль в Чили, финбош в Капской области), что в основном происходит из-за способности к возобновлению путем пневой поросли. С близостью населенных территорий связаны преднамеренные зимние пожары, повторяющиеся через короткие промежутки времени («предписанное выжигание» в Калифорнии). Маквис, будучи «флористическим наследником» третичной лавролистной растительности (см. 15.2.8), наиболее пышно разрастается на влажных участках и на северных склонах. Но, поскольку именно эти участки выбирались под пастбища и пахотные угодья, создается ложное впечатление, что маквис типичен только для сухих скальных и щебнистых мест. Дальнейшая его деградация приводит к формированию открытых низкокустарниковых пустошей — гарриги, или, в восточном Средиземноморье, подушковидных кустарниковых сообществ — фригане (вследствие пожаров, происходящих с интервалом <10 лет или интенсивного выпаса).
С флористической точки зрения растительность областей средиземноморского климата относится к богатейшим на Земле. На ограниченном пространстве здесь произрастают многочисленные озимые однолетники (особенно сем. Asteraceae, Poaceae, Fabaceae), геофиты (в Средиземноморье Orchidaceae, Iridaceae, Liliaсеае), крупные многолетние травы (в Средиземноморье, например. Salvia) и злаки, низкие кустарники (в Средиземноморье Cistus, разнообразные дроки, такие представители Lamiaсеае, как Thymus, Rosmarinus; всевозможные Ericaceae), лианы (в Средиземноморье Asparagus, Smilax), встречаются фрагменты буковых лесов до 10 м высотой — в Средиземноморье с Quercus, Juniperus, Laurus, Pistada (Anacardiaceae), Arbutus (Ericaceae), Rhamnus, Myrtus и дикими Olea', в Калифорнии Quercus, Adenostoma (Rosaceae), Ceanothus (Rhamnaceae), Rhus (Anacardiaceae), Arctostaphylos (Ericaceae); в Чили такие Lauraceae, как Beilschmiedia и Persea и опять же Anacardiaceae; в Капской области и Ericaceae, в Австралии Banksia. Hakea (Proteaceae), а также Mimosaceae (цв. табл. 15.7). На большей части юго-западной и юго-восточной Австралии этот тип растительности представлен эвкалиптовыми рощами с Leptospermum, Callistemon и др. (все — Myrtaceae) (цв. табл. 15.7).
15.2.8. Зона лесов лаврового типа
Зона лесов лаврового типа (лавролистных), которая в третичном периоде была крупным, охватывающим всю Землю биомом, в настоящее время сильно сократилась под воздействием антропогенного фактора и ограничена мелкими реликтовыми разобщенными участками, но тем не менее представленными по всему миру. Она включает вечнозеленые леса, существующие в широких пределах безморозных гумидных областей от северных границ современных субтропиков до умеренной зоны. Находятся они в тех же широтах, что и средиземноморские области, но в районах, где нет выраженного засушливого периода и годовая амплитуда температур относительно невелика. В субтропиках лавролистные леса обычно встречаются в горах, выше 1 400 и до 2 000 м над уровнем моря.
Для зоны лесов лаврового типа типичен климат, приближающийся к гумидному, с количеством осадков 1 000 — 2 000 (до 6000) мм в год. Морозов почти не бывает (зимние минимальные температуры не превышают -2 °С, абсолютный многолетний минимум никогда не превышал -10 °С), в то же время, принимая во внимание распространение биома в пределах почти 25 градусов широты, среднемесячные температуры колеблются от довольно низких значений на севере до высоких в субтропиках.

Сочетание умеренных значений среднегодовых температур и высокого количества осадков соответствуют перегнойным, иногда почти торфянистым почвам с мощной подстилкой из листового опада. Хорошо развит пылевато-суглинистый метаморфический горизонт внутрипочвенного оглинивания. Почвы активно окультуриваются.
Лавролистность — сборное понятие, которое связано не только со специфическими климатическими условиями. Это слово означает характеристику листьев, как «жестких, овальных и цельнокрайних», существующих в выше описанных климатических условиях, причем этот тип листа свойственен в зоне подобного климата только определенному типу лесов. Почти во всех странах света он свойствен также и специфическим видам хвойных пород. Лавролистные леса могут достигать высоты свыше 40 м (прибрежные секвойевые леса Калифорнии, дождевые леса Вальдивии в Чили, эвкалиптовые леса юго-западной оконечности Австралии, леса юго-восточной Азии из Castanopsis и смешанные леса из Nothofagus и Dacrydium юго-запада Новой Зеландии). Часто они представляют собой сохранившиеся реликтовые участки, но все равно не ниже 25 м высоты. Это всегда очень густые насаждения с разреженным подлеском. Холодно-умеренные форпосты этой зоны — дождевые леса подверженных западным ветрам побережий на полуострове Олимпия вблизи Сиэтла (США) и на западе Тасмании (цв. табл. 15.8).
Флористический состав лавролистных лесов представлен, в первую очередь, покрытосеменными, особенно видами сем. Lauraceae и представителями других семейств порядка Laurales, а также семейств Magnoliaceae и Aquifoliaceae (с родом Ilех), которые встречаются в них с высоким постоянством, но низким обилием. Ведущие виды лавролистных лесов юго-восточной Азии относятся к роду Castanopsis, в Новой Зеландии, Тасмании и Чили доминируют вечнозеленые деревья р. Nothofagus (южный бук), в Тасмании и Чили также Eucryphia — представитель собственного примитивного семейства из Rosidae, что подчеркивает древние связи суши южного полушария. В пергумидных климатических зонах южного побережья Австралии доминируют эвкалипты, во Флориде вечнозеленые леса составлены виргинским дубом (Quercus virginiana), но со значительным участием также и Lauraceae (Persea) и Magnoliaceae. Родина цитрусовых — также лавролистные леса юго-восточной Азии; естественные места произрастания этих представителей сем. Rutaceae так и не установлены. Характерные представители хвойных, растущие в условиях этого влажного и мягкого климата — Sequoia sempervirens в Калифорнии, Fitzroya и Araucaria в Чили, виды семейства Podocarpaceae в предгорьях Драконовых гор в Южной Африке и Dacrydium (Podocarpaceae) в Новой Зеландии, Phyllocladus в Тасмании и Cryptomeria в Японии. Лавровые леса на Канарских островах — недавний реликт центрально-европейских. Произрастание лавра благородного (Laurus nobilis) в составе средиземноморского маквиса указывает на его третичное прошлое. В подлеске понтийских лиственных лесов (северная Анатолия) в естественном виде встречаются лавролистные Rhododendron ponticum и Prunus laurocerasus — оба они успешно выращиваются в Западной Европе. Лавролистные горные леса на южных склонах Гималаев (Непал) указывают на древние западно-восточные связи этого биома. В последние десятилетия многие лавролистные виды «убегают» из садов южного Средиземноморья и распространяются в природе. Так, на северном берегу Ладжо Маджиоре (кантон Тесин, Швейцария) уже в наше время под укрытием Castanea сформировался лес лавролистного типа с высокими (до 25 м) камфарными деревьями (Cinnamomum), лавром и веерными пальмами (Trachycarpus из восточной Азии). Rhododendron ponticum довольно давно утвердился на юге Англии и Ирландии, и распространяется там, становясь уже бедствием; последствия внедрения Myrica faya из лавровых лесов Канарских островов в метросидоросовые леса Гавайского архипелага (проникшего вместе с азотфиксирующими симбионтами) уже являются почти катастрофическими.
15.2.9. Листопадные леса умеренной зоны
Типичной растительностью гумидных умеренных широт Северного полушария («неморальная» зона) являются листопадные, летне-зеленые леса; в Южном полушарии есть только маленькие участки лесов этого типа, это листопадные леса из южного бука (Nothofagus). Наиболее высоким видовым разнообразием отличаются такие леса в восточной Азии (Китай, Корея, Япония) и вдоль восточного побережья Северной Америки. Сравнительно бедна в отношении видового состава третья крупная область этих лесов — Средняя Европа (см. 15.11.1), что обусловлено их периодическим вытеснением в ледниковое время, в сочетании с тем, что Альпы и Карпаты при их возвращении играли роль барьера.
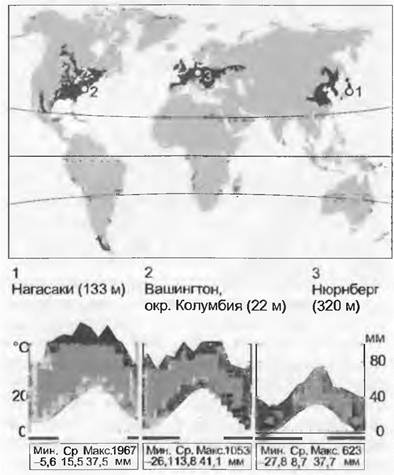
В климатическом отношении равнинные участки этой зоны характеризуются продолжительностью вегетационного периода от 5 до 8 месяцев (при этом в течение 4 — 6 месяцев среднесуточные температуры выше +10 °С), довольно продолжительным холодным периодом (<0°С, с морозами до -25 °С), максимум осадков приходится на лето. Среднегодовые температуры находятся в границах между 5 и 15°С, среднегодовое количество осадков — между 500 и 1 000 мм.
Преобладающий тип почв — слабокислые бурые лесные (камбисоли) с метаморфическими горизонтами разной мощности, формирующиеся, в основном, на лессах. Наиболее развитые и мощные их варианты почти без исключения преобразованы хозяйственной деятельностью, так что современные леса этого типа существуют часто на таких маргинальных вариантах почв, как рендзины или ранкеры. Большая часть лиственных лесов на востоке Северной Америки произрастают сейчас на почвах, которые изъяты из сельскохозяйственного использования 50 — 150 лет тому назад. Ежегодно поставляемый листовой опад полностью разлагается за 1 — 1,5 (2) года, поэтому образование грубого гумуса происходит редко (исключение — кислые почвы под лесами с доминированием Quercus и Castanea), напротив, типично для них Образование гумуса типа мулля или модера.
Прохладно-умеренные лиственные леса в зрелом состоянии достигают высоты 30 — 35 м и относительно светлые (листовой индекс около 5), вследствие чего нижние ярусы их представлены довольно разнообразным набором травянистых растений и кустарников. Произрастают в них также и некоторые вечнозеленые виды, например, встречающиеся почти во всех областях лиственных лесов представители рода Ilex. Флора нижних ярусов большей частью представлена весенне-активными видами (многие геофиты), их жизненный цикл в значительной степени ограничен тем периодом, когда деревья и кустарники еще не разворачивают полностью листву. Исключением являются лиственные леса восточной Азии с бамбуковым подлеском, который настолько плотный, что другие виды произрастать там не могут. Для проростков древесных растений серьезной проблемой выживания становится ежегодное засыпание слоем листьев, в 5 раз превышающем естественную толщину самого листа. Поэтому виды поздних стадий сукцессии имеют гораздо более крупные семена (и сильные всходы). Примерно половина видов этих лесов относится к ветро
опыляемым. Сроки распускания листвы и, в еще более выраженной степени, ее сбрасывания (благодаря чему эти леса переносят морозы и избегают давления снега) регулируются фотопериодизмом, что минимизирует повреждения от поздних и ранних заморозков.
В целом самый важный для этого типа растительности и представленный наибольшим числом видов род — дуб (Quercus). Во всех областях лиственных лесов широко представлены также виды родов Acer, Fagus, Tilia, Betula и Prunus. В восточной Америке произрастает также ряд деревьев из отсутствующих в Европе родов, например, Сагуа (гикори, Juglandaceae, цв. табл. 15.9), Firiodendron (тюльпанное дерево, Magnoliaceae), Liquidambar (Hamamelidaceae), Diospyros (хурма, Ebenaceae). В восточной Азии, прежде всего в умеренных областях северного и среднего Китая, представлены все роды, произрастающие в Европе, но видовой состав их более разнообразен. Многие культивируемые в Европе декоративные деревья и кустарники, например, около 200 видов азалий (летне-зеленые виды рода Rhododendron) происходят из этих восточноазиатских лесов. Хвойные, если присутствуют, представлены почти всегда видами Pinus, в очень сухих районах также видами родов Juniperus и Sabina (например, в северо-восточном Китае). Такие роды, как Aesculus и Platanus, которые в Европе встречаются в естественном состоянии, но только на юго-востоке, также подчеркивают значительную общность этих трех участков зоны лиственных лесов. Сопутствующие прибрежным участкам виды Salix, Alnus и Populus также свойственны всем трем регионам. Восточноазиатская и европейская область соединяются узкой полосой горных лесов, флористически сходных с обеими на родовом уровне и распространенных по южным предгорьям и склонам Гималаев, Гиндукуша и Кавказа (в Непале, к примеру, на высотах между 2 300 и 2 800 м над ур. моря, составленные видами Caprinus, Acer, Betula).
15.2.10. Горные леса умеренной зоны
Наряду с уже рассмотренными тропическими и субтропическими биомами (см. 15.2.2/3) здесь и в следующем разделе будет рассмотрена вторая группа горных биомов (оробиомов), свойственных умеренной зоне (включая высокогорья окраин средиземноморской области). Горные леса умеренного пояса произрастающие в зависимости от географической широты, между 1 000 — 1 500 и 2000 — 3 500 м над уровнем моря, занимают меньшие площади, чем леса равнинных территорий, но зато здесь произрастают флористически чрезвычайно богатые смешанные хвойно-широколиственные леса на нижних уровнях и чистые хвойные леса на высоких.
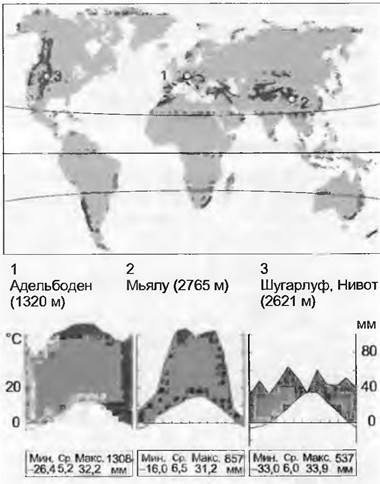
Трудно однозначно дать климатическую характеристику этой зоны, которая представляет собой переход от равнинных лесов умеренной зоны (см.15.2.9, 15.2.12) к умеренно-альпийскому климату, по причине широкого спектра климата— от теплого до умеренно-холодного и от океанического до континентального, а также из-за большой разницы высот, от 1 000 до 2 000 м. В целом в лесах, за небольшим исключением континентальных участков, условия увлажнения достаточно благоприятные, а среднемесячные температуры вегетационного периода ниже, чем на прилегающих равнинах, колеблются между 7 и 12 °С (на равнинах— 12 и 18 °С). Период вегетации длится от 3 до 6 месяцев, зима обычно снежная и холодная (в континентальных районах из-за температурной инверсии не обязательно холоднее, чем в долинах), возможны заморозки в течение 6 — 12 месяцев в зависимости от высоты.
С увеличением высоты почвы становятся все более гумусированными и более кислыми. Общая тенденция — смена по высоте от бурых лесных почв в глубоких долинах до подзолов на больших высотах, причем последние ограничены влажными и умеренно холодными участками. Для верхнего пояса типичны мощные слои подстилки и грубого гумуса.
Эти горные леса необычайно многообразны. Так, в Северной Америке, в Калифорнии в них растут самые крупные деревья в мире, Sequoiadendron giganteum (могучие «мамонтовые» деревья с диаметром ствола до 7 м, высотой 100 м и возрастом 2 000 лет) на высотах ок. 1 500 м над ур. моря; а также вероятно самые старые в мире деревья, Pinus aristata (= Р. longaeva, остистая сосна, 3 500 м над ур. моря). Но наличие возобновления из частей ствола или у некоторых видов из пневой поросли, т.е. однозначно из того же генетического материала, делает вопрос о «самых старых» деревьях спорным. В горах всех областей Голарктики существуют леса с доминированием Abies, Picea и Pinus. На северо-западе США (например, г. Маунт-Райнер) и местами в гумидных горах северовосточной Азии на границе лесного пояса (например, г. Фудзи) доминируют виды рода Tsuga,вымершего в Европе во время оледенения. В сухих зонах к ним присоединяются горные леса из Juniperus и Cupressus (Каскадные горы, Атлас и высокие участки гор Средиземноморья, Каракорум, Тибет). В умеренно-горных поясах южного полушария хвойные редки (Phyllocladus и Arthrotaxis в Тасмании, предствители рода Podocarpus в юго-восточной Австралии, Austrocedms в Чили). Лиственные породы в голарктических горных лесах представлены родами Betula, Sorbus, Ainus и Populus, в Азии также Crataegus, в южном полушарии в субантарктической зоне — представителями родами Nothofagus (как вечнозелеными, так и листопадными), в Австралии морозостойкими видами Eucalyptus. К самым высоким деревьям (> 110 м) Земли относится Eucalyptus regnans («горный ясень»), произрастающий на горных склонах по южному краю Снежных гор (цв. табл. 15.10).
Горные леса чрезвычайно важны для защиты лежащих ниже районов, так как они препятствуют эрозии, а в очень снежных районах предотвращают сход лавин. Во многих местах верхняя граница леса отступает под воздействием рубок и освобождения площадей под пастбища (см. 15.2.2). Своей естественной границы распространения горные леса умеренного пояса по всей Земле достигают на уровне, где среднегодовые температуры становятся ниже 6 — 7 °С (исключения: склоны с рыхлым сыпучим субстратом, лавинные полосы, или области с крайне океаническим климатом, где границы лесной зоны располагаются ниже). Ввиду того, что зимние температуры на границе леса по всему миру очень сильно варьируют, их значения не играют столь большой роли. По другим климатическим факторам верхнюю границу горных лесов определить легче: она проходит на расстоянии + 100 м по высоте от сезонной изотермы (лета) 6 — 7°С. Жизненная форма дерева тесно связана с макроклиматом, поэтому предположительно в более низких температурных границах могут существовать только более низкорослые растения, которые создают свой собственный (более теплый) микроклимат.
15.2.11. Альпийская растительность высокогорий умеренной зоны
Несмотря на то что высокогорья, находящиеся выше естественной границы леса (альпийский оробиом), занимают только около 3 % земной поверхности, их флора составляет около 4 % от всей флоры цветковых растений. Крупные горные системы умеренной зоны — это Скалистые горы и Сьерра-Невада в Калифорнии, Каскадные горы, Альпы, Карпаты, Кавказ, северная часть Гиндукуша и Гималаев и их продолжение вплоть до Кореи, горы Центральной Азии, Японии, Анды южнее 35°, Драконовы горы в Южной Африке, Снежные горы в Австралии, Крэйдл в Тасмании и Новозеландские Альпы, в сумме это почти половина альпийских территорий Земли. Общепринятый термин «альпийский» в ботанико-географическом понимании означает «расположенный выше границы леса», что отличает его от более широкого понимания в смысле «горный».
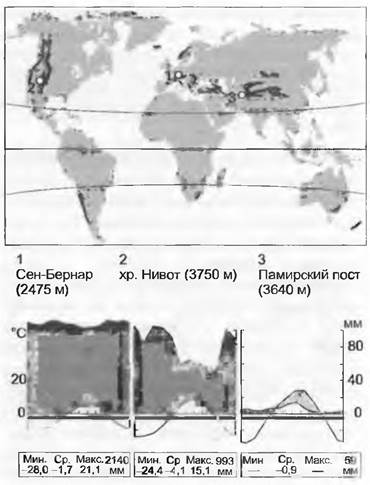
Средние значения температур в припочвенном слое воздуха или в области подземных меристем в течение короткого (от 6 до 16 недель) периода вегетации лежат в границах от 5 до 10 °С, в полдень при солнечной погоде они поднимаются до 20 °С. Данными метеостанций не отображаются микроклиматические ситуации, играющие существенную роль. Благодаря своему низкому росту и компактным жизненным формам растения в течение вегетационного периода, по крайней мере в дневные часы, избегают холода (морфологические приспособления для улавливания солнечного тепла). Для тех мест, где проводились тщательные повторные измерения, доказано, что предполагаемый обычно дефицит влаги для альпийских растений совершенно несуществен; может быть он есть только в очень сухих районах Памира и в части Южных Анд (локальное исключение — скальные кромки с тонким слоем мелкозема). Данные по сумме осадков (так же, как и по среднегодовым температурам) дают мало информации о фактических условиях жизни в этом поясе, поскольку она в течение длительного времени неактивна из-за холодных условий. Благодаря таянию снегов и летним дождям в течение всего короткого периода роста в распоряжение растений поступает почти вся годовая сумма осадков.
В нижней части альпийского пояса (особенно при наличии кустарничков и дерновинных злаков) благодаря пониженной скорости разложения преобладают перегнойные почвы с очень мощным слоем грубого гумуса. Поверхностное иссушение почв может периодически тормозить процессы минерализации и затруднять доступность питательных веществ. С увеличением высоты все чаще встречаются неструктурированные примитивные почвы. Важное значение приобретают криогенные процессы и механическая эрозия почв, обусловленная крутизной склонов. Под щебнем или обломочным материалом скрывается иногда неожиданно много влаги. Стабильность почв и при этом защищенность более низкорасположенных районов существенно зависят от растительного покрова альпийского пояса.
Безлесная по определению альпийская растительность (здесь объединяются собственно альпийский и субальпийский пояса) состоит главным образом из (1) кустарничков, (2) образующих вегетативные клоны граминоидов (злаки и осоки), (3) многолетних, часто также клонообразующих, формирующих розетки травянистых растений, (4) подушковидных растений в широком смысле (образующих коврики, плоские подушки, полушаровидные подушки) и (5) криптогамных растений (мхов и лишайников). Наибольшего разнообразия достигают розеточные растения и криптогамы (цв. табл. 15.11). Очень мало или вообще нет геофитов и однолетников (за исключением районов, пограничных с субтропиками или Средиземноморской областью). Морфологические, фенологические и физиологические адаптации настолько сильно развиты, что продуктивность сомкнутой альпийской растительности, пересчитанная на месяц (!) активного роста, соответствует всем остальным гумидным биомам, включая равнинные тропики (см. рис. 13.39). Лишь одно значимое ограничение здесь — длительность периода активного роста, поэтому растения, формирующие этот биом, никоим образом не более угнетены, чем в других биомах. Морозы, приходящиеся на период зимнего покоя, не играют существенной роли. Критическое значение имеют только раннелетние поздние заморозки и ранние осенние морозы, при этом они никогда не представляют собой существенной угрозы, будучи причиной только частичной потери листьев и опадения семян. Очень распространено вегетативное РАЗМНОЖЕНИЕ.
В умеренной зоне в альпийском поясе наиболее значимые семейства — Asteraceae, Роасеае, Сурегасеае, Caryophyllaceae, Ericaceae, Gentianaceae, Rosaceae и Ranunculaceae. Первые четыре обычно составляют более 50% локальных флор. В разных регионах существенна роль Saxifragaceae, Primulасеае, Campanulасеае, Роlygonaceae и Scrophulariасеае. Высокогорья, подобно островам в океане, в значительной мере изолированы друг от друга, поэтому они богаты локальными эндемиками. Флора Альп объединяет около 650 видов цветковых растений с альпийским типом распространения, около 150 видов поднимается выше 3 000 м. Мировой рекорд принадлежит Saussurea gnaphalordes, растущей на высоте 6 400 м в массиве Эвереста.
15.2.12. Степи и прерии
В континентальных областях умеренной зоны образуются обширные злаковники, причем помимо климата эти экосистемы формируются под влиянием таких регионально проявляющихся факторов, как копытные звери и пожары, отчасти также и человек. Кроме евроазиатских степей и североамериканских прерий (и растительности Большого Бассейна) аналогичные формы растительности есть также в Аргентине (пампа), на умеренно-теплом юге Африки и в горах Атласа, фрагментами — в умеренной Австралии и на подветренной стороне Новой Зеландии, в горах Южного острова.
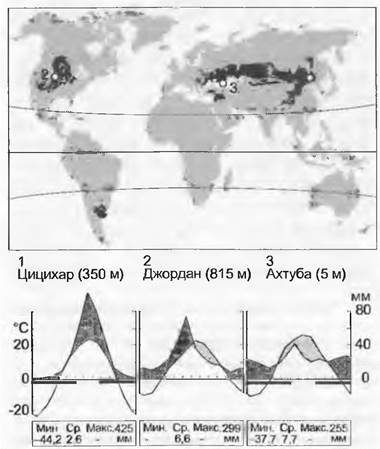
Среднегодовые климатические данные (средние суммы) очень мало могут сказать об условиях жизни на территории этого биома. Во внутренних частях крупных материковых масс северного полушария между 35 и 55° широты зимы отчасти экстремально холодные (до -50 °С), лето же экстремально жаркое (часто > 40 °С). Запасы влаги в почве в течение вегетационного периода складываются из тающего зимнего снега, из дождей, преобладающих в весеннее время, и, в более влажных районах, из летних грозовых дождей (годовая сумма осадков колеблется в пределах между 250 и 500 мм).
Почвы развиваются в основном на лёссах, лёссовидных суглинках или песках, и часто имеют очень глубокий профиль. Так как большая часть первичной продукции растений остается под землей (в злаковниках это 2/3), формируется глубокий гумусовый профиль, часто темноокрашенный (черные земли, черноземы). Роющие грызуны (хомяки, луговые собачки) содействуют перемешиванию почвенных слоев.
Грубо говоря, в этой климатической зоне различают четыре типа растительности: (1) злаковые сообщества высотой ниже 50 см — это низкотравные прерии, типичные степи, нагорные степи Монголии (сухие и холодные районы); (2) злаковые сообщества выше 1 м, из мощных дерновинных злаков, высокотравные прерии, пампа, южноафриканские «прерии» (более влажные области с более мягким климатом); (3) полынные степи, где доминируют одревесневшие у основания или полностью полукустарники Artemisia (особенно холодные области в Большом Бассейне и Центральной Азии); на плоскогорьях Средней и Передней Азии они сменяются (4) колючеподушечниками (Astragalus, Acantholimon, Noaea)(цв. табл. 15.12). Многие из этих континентальных сухих областей лежат довольно высоко, чем объясняются низкие температуры воздуха, несмотря на довольно южные широты). Низкотравные прерии Вайоминга и Монтаны, а также Большого Бассейна лежат на высоте ок. 2000 м, монгольские степи — на 2 500 м, в умеренной области Южной Африки (к западу от Драконовых гор) на высоте 1 600 м. Благодаря тому, что хорошие почвенные и климатические условия благоприятны для возделывания зерновых культур и пастбищного животноводства («Дикий Запад»), естественные степи и прерии сейчас почти совершенно исчезли. Но выпас копытных животных (например, стад бизонов) был также и естественным фактором нагрузки. Части континентальных злаковников могли бы быть по условиям климата и облесены (районы с годовой суммой осадков >400 мм), но там росту деревьев препятствовали, как и в саваннах, естественные пожары, возникшие от молний. Ростовая меристема злаков защищена многими сантиметрами лежащей выше нее земли. Первоначальное влияние копытных здесь не так существенно, как в саваннах. Дерновинные злаки, такие, как в высокотравных прериях и в пампе, могут также препятствовать росту деревьев как сильные конкуренты.
Характерный род злаковых степей, отчасти также низкотравных прерий — Stipa (ковыль). Stipa еще произрастает в настоящее время в сухих местоположениях Центральной Европы (например, Кайзерштуль, Валлис, Винтшгау, Вахау), являясь реликтом холодных и бедных осадками приледниковых окраин периодов оледенений, его западный предел распространения лежит в Паннонской области. Stipa tenacissima (ковыль альфа) доминирует в степях Атласского нагорья. В низкотравных прериях важную роль играет Boutelouagracilis (англ. «Blue Grama»). В американских высокотравных прериях доминируют С4-злаки, относящиеся к родам Andropogon, Sorghastrum и др. (средний юг Северной Америки, для Канзаса типично обилие таких очень декоративных двудольных растений, как Echinacea и Rudbeckia), Hypatrhenia и Pennisetum доминируют в Южной Африке, a Cortaderia в аргентинской пампе. На большей части запада Северной Америки преобладает Artemisia tridentata; А. sieberi сменяет злаки в сухих частях Центральной Азии. Континентальная засушливая область Передней Азии является также родиной пшеницы, ячменя и ржи, южные предгорные области центрально-азиатских степей — фруктовых деревьев сем. розоцветных (яблоня, абрикос; Алматы — город яблок).
15.2.13. Пустыни умеренной зоны
В умеренной зоне также имеются пустыни и полупустыни. Зимы морозные, а летом осадков очень мало, как правило, это еще осложняется сильным засолением почвы, что создает крайне экстремальные условия существования на тех же широтах, на которых в условиях океанического климата формируются буковые леса. Границы с субтропическими засушливыми областями очень постепенны. Так, сюда относится пустыня Мохаве, в которой благодаря положению в рельефе зимы очень холодные. Несмотря на то что пустыня находится на краю Долины Смерти в северо-восточной Калифорнии, лежащей в депрессии, ниже уровня моря, в летнее время одного из самых жарких мест Земли, она относится к пустыням умеренного, зимне-холодного типа. Растительность вокруг Каспийского моря соответствует растительности вокруг всех крупных соленых озер. Умеренные полупустыни есть на низменностях Центральной Азии, в Гоби и некоторых внутренних долинах г. Гиндукуш, в Южном полушарии — в Патагонии и частично в южной Австралии (цв. табл. 15.13).

Климат этой зоны очень сходен по температурному режиму с описанными выше злаковниками (см. 15.2 12), правда, с более высокими максимальными температурами в летнее время, но осадков меньше 250 мм (в пустыне Так- ла-Макан в Центральной Азии — менее 60 мм, в пустыне Гоби — менее 100 мм). Почвы, как и в остальных засушливых областях, слабо развитые. Там, где воздействует засоление, формируются экстремально щелочные почвы — солончаки (соленые или гипсовые почвы) и солонцы (содовые почвы, см. 12.5.2.3). При высоком содержании гипса в почве могут образовываться твердые, почти непроницаемые для корней растений, водоупорные слои. Избыточное засоление или загипсовывание всегда возникает тогда, когда длительное испарение (происходящее путем поднятия влаги из почвы) превышает количество осадков.
Растительность этой семиаридно-аридной области с холодными зимами меняется от сравнительно разнообразных сочетаний видов и жизненных форм, как в пустыне Мохаве, или также эдафически обусловленных (песок) островных пустынях южной Австралии, до суккулентных галофитов, образующих одновидовые сообщества. За небольшими исключениями (например, на скатах), всегда значительную роль играет почвенное засоление. По площади на большей части этой зоны доминируют галофиты. Мохаве широко известна по ее полусуккулентным «хохлатым» канделяброобразным деревьям юкки (Yucca brevifolia), но здесь также богато представлены морозоустойчивые виды р. Opuntia, кустарнички и однолетние злаки и разнотравье. На плато, окружающих центральную часть Долины Смерти с ее раскаляющимися в летнее время почвами (абсолютный максимум 56 °С в тени, ежедневно температура поднимается выше 45 °С), доминируют кустарники Atriplex (их листья покрыты солевыми выделениями, поэтому они белые и хорошо отражают тепло). По той же причине у многих видов на листьях развито беловойлочное опушение (например, у сложноцветного Encelia farinosa). Как и на морских побережьях, растительность, окружающая засоленные впадины, в разной степени устойчива к засолению в зависимости от удаления от центра. Так, вокруг больших соленых озер обилие умеренно толерантных к засолению кустарничков и травянистых многолетников, относящихся в основном к семейству Asteraceae (Юта; такая же ситуация вокруг Каспийского и Аральского морей), постепенно снижается на открыто залегающей кристаллической соли, пожалуй, наиболее неблагоприятном для жизни биотопе Земного шара, растет только один вид, Suaeda depressa (для него характерен CAM-газообмен, активное выделение солей, осмотическое давление клеточного сока до 7 мПа). Почвы гипсовых пустынь, таких, как центрально-казахстанские, химически не так агрессивны (хотя pH 11), но зато здесь имеются механические препятствия, ограничивающие доступ к грунтовым водам. Лишь немногим растениям изредка удается пробить лежащий на глубине 1 — 2 м водоупорный слой. Растительность этой зоны состоит преимущественно из частично одревесневших кустарников (но саксауловые «леса» у оз. Балхаш достигают 12 м высоты, хотя обычно они не выше 3 м). Центрально-европейский форпост галофильной растительности засоленных степей находится на Паннонской низменной равнине (например, на восточном берегу оз. Нойзидлер-Зее).
Во флористическом отношении это самая простая часть зонобиома, так как повсеместно доминирует одно семейство, а именно Сhenopodiасеае (Аtriplex, Suaedа, Salicornia и др.). В отдельных регионах играют роли также представители Zygophyllaceae.Solanасеае, Polygonасеае и Asterасеае. За пределами засоленных областей флора богаче и в более теплых районах видовой спектр приближается к субтропическим засушливым областям (см. 15.2.6).
15.2.14. Бореальные леса
Бореальная («северная») часть умеренной зоны охватывает широкий пояс всей Голарктики, где доминируют леса из хвойных пород («тайга»). Они захватывают лесную область северной Европы (Скандинавия, северная часть России), Сибирь, Канаду и Аляску, достигая Полярного круга и местами заходя за него (в Восточной Сибири почти до 73° с. ш.).

В этой зоне влажность также сильно меняется по градиенту от морского побережья до внутриконтинентальных районов, но из-за короткого вегетационного периода (3 — 5 месяцев) и в общем низких температур она редко является здесь лимитирующим фактором. Часто здесь наблюдаются очень низкие зимние температуры (в Сибири до -70 °С), но лето может быть жарким и даже знойным.
Почвы варьируют от богатых перегноем бурых лесных почв и подзолов (светлоокрашенные почвы с горизонтом вымывания) до болотных почв и слаборазвитых почв типа ранке - ров и рендзин. Примитивные почвы на песках также зарастают лесом. Медленно разлагающийся опад хвои способствует кислой реакции почвы. При наличии в почве многолетней мералоты развитие сомкнутых лесных насаждений исключается, причем между почвенной температурой и лесом существует обратная связь. Лес, смыкаясь при таянии мерзлоты, способствует, в свою очередь, понижению температуры почвы (солнечное излучение, приносящее тепло, больше не достигает почвы) и многолетняя мерзлота вновь поднимается, деревья отмирают, при этом Солнце опять начинает нагревать почву сильнее и так далее. Незначительное, мощностью в несколько сантиметров, превышение уровня воды может быть решающим фактором становления леса. Из-за этого даже небольшие колебания рельефа глазомерно кажутся значительными (в переувлажненных стоячими водами низинах лес не растет).
Сильные зимние морозы исключают произрастание в бореальных лесах большинства лиственных пород. Но примечательно то, что именно в экстремально холодных районах доминирует летнезеленая лиственница, иногда образующая одновидовые насаждения. Типичный бореальный лес более открыт, чем леса умеренной зоны, так как в большинстве случаев он еще не нарушен, в нем много сухостоя, отмершие деревья сохраняются на корню десятилетиями. В естественных циклах развития леса большую роль играют периодические (раз в 200 — 300 лет) пожары. В последние десятилетия частота пожаров в канадской тайге заметно возросла в связи с общим потеплением климата. С приближением к северной границе деревья становятся более сбежистыми, а интервалы между ними увеличиваются. Собственно, на границе леса в отдельных регионах хвойные леса сменяются низкорослыми березовыми (березовая лесотундра). Хвойные деревья часто ветвятся на уровне почвы и истончаются, что при низком стоянии Солнца и преобладающей редкостойности насаждений способствует более полному использованию света, снежный покров становится тоньше, что положительно влияет на нагревание почвы (и при этом на корни деревьев). Плотный моховой покров под деревьями может задерживать Питательные вещества и лес начинает «голодать» (влияние пожаров в этом случае благоприятно). Питанию деревьев способствует развитее эктомикоризы (со шляпочными грибами). Появляющийся подрост местами подвергается зимой сильному прессу со стороны животных. Благодаря широкому распространению эта лесная область является важным климатическим фактором, так как из-за низкого альбедо (отражательной способности), она абсорбирует больше солнечной радиации, чем безлесные, белые в зимнее время пространства. К тому же она накапливает в древесине и гумус- ном слое очень много углерода; 40 % всей древесины этих лесов используется для производства бумаги, поэтому они относятся к остро угрожаемым.
Доминирующие роды в бореальных лесах — Picea, Pinus, Abies, в отдельных районах также Larix (цв. табл. 15.14). На западе северной Скандинавии доминирует Pinus sylvestris, но уже в восточной Финляндии начинается область распространения Picea obovata (подвид Р. abies, с мелкими шишками). В Северной Америке такую же роль играет Picea glauca. Abiesbalsamea в Северной Америке соответствует A. sibirica в Сибири. При неблагоприятных почвенных условиях в целом преуспевает Pinus. Larix доминирует прежде всего в Восточной Сибири («светлохвойная» тайга из L. dahurica и др., в Северной Америке ее заменяет L. laricina). Важнейшие роды, представляющие лиственные деревья-спутники бореальной тайги — Betula, Populus (Р. trémula, европейская осина; Р. tremuloides, американская осина), Salix, Sorbus. В нижних напочвенных ярусах, кроме мхов и лишайников, доминируют кустарнички, в основном род Vaccinium.
15.2.15. Субарктическая и арктическая растительность
В то время как в Антарктиде обнаружено только 2 коренных вида сосудистых растений и площадь, свободная ото льда, очень мала, циркумполярный ботанико-географический пояс Северного полушария, расположенный к северу от границы леса, занимает 5 % площади земной поверхности и в нем произрастает почти 1 000 видов покрытосеменных. Сомкнутый растительный покров Субарктики называется тундрой (между 62 и 75° с. ш., в Европе и Гренландии благодаря влиянию Гольфстрима она продвигается на 5 — 7° севернее, чем на востоке Северной Америки). Это обозначение взято от финского слова «tunturi», обозначающего безлесные холмистые местности. В более высоких широтах растительность сильно фрагментирована и приурочена к благоприятным местообитаниям (до 83° с. ш.).
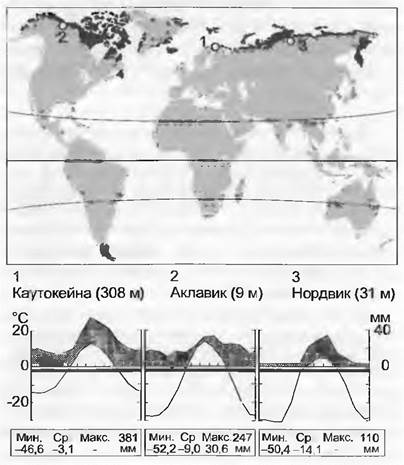
Климат арктического биома характеризуется коротким, длящимся 6 —16 недель вегетационным периодом, большая часть которого приходится на 24-часовой полярный день. В это время температура в южной части тундры может подниматься до 20 °С. Образующийся в течение длинной зимы снежный покров очень маломощный, поскольку общее годовое количество осадков низкое (<400 мм), следствием чего является глубокое проникновение в почву арктического холода. Несмотря на малую сумму осадков арктический биом (за небольшими исключениями) сырой, так как испарение очень низкое. Кроме того, многолетняя мерзлота во многих местах препятствует просачиванию влаги в почву (см. ниже). Локальное перераспределение влаги сильно связано со снежным переносом (рельефом).
Холод и застойное увлажнение и обусловленное ими торможение процессов разложения накладывают отпечаток на арктические почвы. Они часто заболочены, очень кислые, на них могут селиться только специализированные растения (Ericaceae, Сурегасеае). Структура почвенного покрова формируется под влиянием процессов промерзания-протаивания (солифлюкция, полигональные почвы, морозное пучение). Растительную мозаику определяют рельеф и связанное с ним застойное увлажнение. Хорошо дренированные почвы на склоновых участках обусловливают увеличение здесь числа видов и продуктивности. Там, где летом почвы глубоко оттаивают, развит соответственный растительный покров.
Основные формы арктической растительности таковы: (1) кустарничковая тундра, (2) осоковая и пушицевая тундра, (3) болота, (4) открытые сообщества высших растений на примитивных почвах, (5) моховая и лишайниковая растительность (дв. табл. 15 15). На южной границе тундры в отдельных районах есть редкостойная березовая (или лиственничная — Ред.) лесотундра, которая переходит в бореальные леса. Почти все растения этого биома способны к вегетативному распространению (образованию клонов). Повсеместно в растительном покрове существенную роль играют споровые растения (мхи и лишайники), образуя напочвенный ярус. В высокой Арктике примитивные почвы покрыты коркой из криптогамных растений с высоким участием сине-зеленых водорослей. Большую роль в снабжении растений питательными веществами играют ассоциирование с почвенными микроорганизмами (осоки) и Симбиоз с грибами (микориза вересковых). В почвах, преимущественно кислых, помимо аммонийного азота источником последнего служат также свободные Аминокислоты. Наличие долгого светового дня в какой-то степени компенсирует краткость вегетационного периода. Поскольку феноритмика растений регулируется в значительной степени фото периодически, сама по себе теплая погода не может заменить растениям фактического срока освещения в течение года. Многочисленные исследования, проведенные в альпийском поясе, показали, что низкие температуры во время вегетационного периода не ограничивают роста, как часто полагают. Ограничивающий фактор — длительность этого периода.
Флора субарктической и арктической зон относительно бедна (<1/10 от видового разнообразия всей альпийской флоры). Это разнообразие в основном сосредоточено на небольших по площади участках, определяемых рельефом, на которых нет застойного увлажнения. Когда поверхность плоская и притом еще мокрая, видовое разнообразие сильно снижается, так что большая часть первичной продукции формируется менее, чем 100 видами. Наибольшую роль играют 3 семейства: Ericaceae (в особенности роды Vaccinium, Empetrum), Сурегасеае (Carex, Eriophorurri), Salicaceae (Salix). Другие важные семейства — Betulaсеае (Betula папа), Rosaceae (Rubus) и Роасеае (Deschampsia и др.). Важнейшие роды мхов — Sphagnum (торфяной мох) и Hylocomium (этажный мох), широко распространены кустистые лишайники, относящиеся к родам Cladina, Cladonia и Cetraria. Арктоальпийская флора обнаруживает большое сходство с флорой альпийского пояса умеренной зоны (общие виды, как Ranunculus glacial is и Oxyria digyna), однако все же неуместно употреблять понятие «тундра» по отношению к альпийской растительности.
15.2.16. Растительность морских побережий
Растительность морских побережий носит как зональные, так и азональные черты. Зональные, типичные для данного климата формы в свойственных ему широтах распространены существенно шире, чем остальные зональные типы растительности. Климат очень ровный. Растения прибрежных местообитаний подвержены существенно меньшим амплитудам температур, зимы в очень широких границах умеренной зоны почти всегда безморозные.
Во всех случаях эта растительность находится под воздействием засоления и механических факторов (ветер, затопление, нестабильность почв). Растения крутых берегов должны быть крайне солеустойчивыми, находясь в зоне постоянного воздействия брызг морской воды, так как после их испарения остаются чистые отложения соли. Песчаные дюны, казалось бы, сильно нагреваемые и крайне сухие, на самом деле часто вовсе не сухие. Крупнопористый песок препятствует капиллярному поднятию влаги, в результате чего она консервируется в более глубоких слоях, которых корни растений успешно достигают. Возможное из-за этих процессов сильное транспирационное выхолаживание и собственное затенение растениями препятствуют сильному нагреванию поверхности песка и связанным с этим повреждением. От перегревания листьев на таких сильно нагреваемых солнцем местообитаниях многие растения защищаются (1) наличием очень узких листьев (хороший термический обмен с воздухом) (2) отражающей верхней поверхностью (опушение шерстистыми или щитовидными волосками, см. рис. 13.10) или (3) направленными вверх пластинками листьев, следующими за положением Солнца, как, например, у широко распространенного в настоящее время Hydrocotyle bonariensis (см. ниже). Растения илисто-глинистых наносов должны быть приспособлены к тому, что их корни будут существовать в условиях отсутствия кислорода (анаэробных), что в совокупности с непосредственным воздействием соленой морской воды при сильном солнечном свете приводит к выработке разнообразных приспособлений, которые свойственны растительности мангров (см. ниже).
В общем все обитающие на побережьях растения подвержены механическому воздействию со стороны штормов, прибоя и / или нестабильного грунта. Это объясняет почему здесь так часто встречаются клонообразующие жизненные формы и устойчивые к сильным ветрам листья. Семена и плоды также имеют специальные приспособления, что наглядно видно из строения кокосового ореха. Внутри трехслойной защитной оболочки твердого плода: кожистой (экзокарпия), волокнистой плавучей (мезокарпия) и непромокаемой твердой (эндокарпия) находится крупное семя с богатым жирами, водонепроницаемым эндоспермом (копра) и жидкой питательной средой внутри (плавающий эндосперм), который способствует прорастанию зародыша этой плавающей диаспоры после тысячекилометровых странствий по океану. Зародыши Rhizophora (мангровые деревья) способны укрепляться на подвижных илисто-глинистых берегах благодаря своему якореобразному корню и удлиненному гипокотилю.
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.