БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
14. ЭКОЛОГИЯ ПОПУЛЯЦИЙ И РАСТИТЕЛЬНЫХ СООБЩЕСТВ
14.2. Ареалы растений
Ареалы растений — это области распространения вида или более высоких таксонов. Они представляют собой результат исторического и пространственно-временного распространения или отступания таксона (рис. 14.14, см. 10.3). Данные ареалы определяются морфологической и экофизиологической конституцией (в соответствии с приспособляемостью), конкурентной силой, возможностями распространения на протяжении всей истории Земли и произрастанием в пригодных для этого местообитаниях. Даже активно расселяющиеся таксоны далеко не всегда занимают все возможное для себя жизненное пространство («потенциальный» ареал в противоположность фактическому), поскольку перемещения и стойкое заселение происходят медленно или расселению препятствуют ограничивающие факторы (например, моря, горы или массивы пустынь). Крайне быстрое или, напротив, продолжительное распространение многих видов, расселяющихся с помощью человека (см. антропохория), показывает это наиболее выразительным образом. При возникновении современных ареалов в игру вступают также генетические, экологические и исторические факторы. Наука об ареалах описывает и сравнивает ареалы распространения таксонов (большей частью видов, а также родов или групп близкородственных видов). На основе этого могут быть выяснены комплексные связи между формами ареалов и условиями существования В настоящее время и в прошлом.
Рис. 14.14. Происхождение типов ареалов растений (распространение — по горизонтали, время — по вертикали снизу-вверх, современное состояние — на срезе ровной поверхности, вымершие популяции — ниже уровня этой поверхности): А — расширение ареала (например, Trifolium repens, см. рис. 10.26). Отмирание популяций и сокращение до дизъюнктивного ареала В (например, Pinus nigra, рис. 10.31) или до реликтового палеоэндемичного С (например, Gingko biloba, см. рис. 14.16); D — аллопатрическая дифференциация близкородственной группы на три викарирующих таксона (например, Erysimum sect. Cheiranthus, продукты аллопатрического образования родственных видов, так называемых схизоэндемиков различных участков Эгейской области); Е — псевдовикарирование двух не близкородственных, но экологически и, соответственно, географически замещающих друг друга видов (например, Gentiana clusii и G. acaulis s. str. (= G. kochiana); F — круг форм с единым центром многообразия (Z — центр формирования, R — реликтовые эндемики, N — неоэндемики) (например, Carlina, см. рис. 14.21). Схема поясняет, что между возрастом таксона, разнообразием его форм и размером его ареала отсутствует непосредственная связь

14.2.1. Типы ареалов
Ареалы составляются путем объединения местонахождений одного таксона и лучше всего отражаются на картах ареалов (рис. 14.15. см. рис. 10.31, 11.259, 14.16 — 14.18). Содержательность изображения ареала зависит от правильного систематического ограничения таксона или его отчленения от группы близкородственных видов, а также от достоверности флористических данных о его точках произрастания. Даже для ареалогического анализа флоры сосудистых растений Центральной Европы нет еще пока полной систематической и флористической основы.
Картографическое представление о распространении таксонов нуждается в некотором абстрагировании, поскольку частота встречаемости и пространственное размещение индивидумов в мелких и более крупных местах обитания с учетом находящихся между ними промежутков обычно очень неравномерны Широко распространены точечные и контурные карты, или оба эти метода комбинируются (см. рис. 10.31, 14.25). При современных ареалогических исследованиях с применением электронных методов обработки данных (например, при проведении международных работ по картированию флоры Центральной Европы) особенно хорошо показали себя растровые карты, на которых наличие или отсутствие таксона в каждом случае наносилось на поле картирования определенного фиксированного размера (см. рис. 14.15). На высотных профилях можно также представлять вертикальное распространение определенного таксона.
Для описания, сравнения и анализа ареалов таксонов важны следующие критерии:
— размер: от локального (эндемичного) до континентального и +- распространенного по всему миру (космополитного);
— непрерывность: от полностью замкнутого (непрерывного) до сильно фрагментированного (дизъюнктивного);
— плотность заселенности: повсеместно, широко, рассеянно или редко;
— распределение многообразия форм таксона в пределах ареала;
— положение по отношению к ареалам близкородственных таксонов;
— географическое положение ареала.
Путем сравнительной оценки многочисленных ареалов с этих точек зрения могут быть установлены различные типы ареалов (см. рис. 14.14).
Рис. 14.15. Пример растровой карты (предварительной) из проекта «Картирование флоры Центральной Европы»: синеголовник полевой (Eryngium campestre, Apiaceae).
Представлено присутствие или отсутствие вида в полях градусной сетки длиной 10x6' географической долготы и соответственно шириной примерно 12x6 км. Субсредиземноморско-понтийский вид распространяется по теплым и сухим логам и долинам рек далеко на север

14.2.1.1. Протяженность ареалов
Между таксонами, произрастающими только в одном местонахождении в мире, и теми, которые в пределах своей приуроченности к определенному местообитанию произрастают по всему миру, имеются все возможные промежуточные формы. Таксоны, ограниченно произрастающие в пределах единственного, как правило, небольшого ареала, называются эндемичными, причем это понятие используется довольно неопределенно. Наряду с действительными редкостями, в основном на видовом уровне локальными эндемиками,различают также региональные (например, произрастающие только в Альпах) или даже континентальные эндемики (например,
произрастающие только в Австралии), причем последняя категория имеет смысл только на уровне семейства. Есть эндемики старые, реликтовые — палеоэндемики и молодые — неоэндемики. Широко распространенные космополиты — это преимущественно растения, произрастающие в условиях, связанных с деятельностью человека, но есть и природные космополиты, прежде всего среди споровых растений.
Из известных реликтовых эндемиков, которые когда-то были распространены широко, можно назвать Ginkgo biloba (сейчас он еще растет только в западной части Китая; рис. 14.16), Sequoiadendron giganteum (Калифорния) и Welwitschia mirabilis (юго-западная Африка). В противоположность им эндемичными ареалами обладают Betula oycoviensis (юго-восточная Польша), Papaver kerneri (юго-восточные Доломитовые Альпы); виды рода Erysimum sect. Cheinmthus в Эгейской области (см. рис. 10.32), как известно, возникшие в результате скрещивания, — это расположенные рядом ареалы близкородственных видов, все они относятся к относительно молодым эндемикам (новым видам). Доля эндемичных таксонов, очевидно, увеличивается с возрастом и степенью изоляции жизненного пространства.
Рис. 14.16. Распространение рода Ginkgo в Северном полушарии по ископаемым находкам от нижней юры до раннего плиоцен-неогенового периода и нашего времени.
В основе карт лежат современные очертания земной поверхности. Изменения с начала юрского периода (почти 200 млн лет назад) оказывали незначительное влияние на потенциальные пути миграции в Северном полушарии

В качестве примеров космополитных видов среди споровых растений можно привести печеночный мох Marchantia polymorpha или папоротник Ptendium aquilinum, болотные и водные растения, широко распространяющиеся водными птицами: водяную сосенку (Hippuris vulgaris), тростник (Phragmites australis), а также многочисленные, распространяемые человеком сорняки, которые на нарушенных участках (рудеральных местах, часто у обочин дорог) можно найти по всему земному шару (например, вилы родов Plantago, Роа, Rumex, Senecio, Stella ria. Trifolium). На уровне семейств по всему миру встречаются те, которые по своей природе имеют высокое видовое богатство (Orchidaceae, Роасеае, Asteraceae, Fabaceae).
14.2.1.2. Естественные разрывы ареалов
Полностью непрерывное заселение встречается редко. По меньшей мере по направлению к окраинам ареалов распространение большинства таксонов рассредоточивается, ограничиваясь отдельными форпостами (или, наоборот, местонахождениями, оставшимися при отходе). Если пробелы в ареалах оказываются такими большими, что таксоны не могут их преодолеть обычными способами распространения, говорят об эксклавах и, соответственно, дизъюнкциях. Многие дизъюнктивные узоры повторяются довольно регулярно (генетические последствия искусственного фрагментирования местообитаний, см. 10.3.2.2).
У Pinus sylvestris на севере мы видим относительно сплошной основной ареал, в южных же горах имеются дизъюнктивные, более мелкие частичные ареалы, или эксклавы. Почти одинаковы по протяженности три дизъюнктивных частичных ареала рода Fagus (рис. 14.17). Тот факт, что дизъюнкции у Fagus, Hepatica (см. рис. 14.17) и других родов, свойственных широколиственным лесам, очень сходны, объясняется общими историческими и климатическими причинами. При наличии дизъюнкций можно думать или о редукции сплошной в прошлом области распространения или, в чрезвычайных случаях, о привнесении из очень далеко расположенной области. Для гибридогенных таксонов (например, у аллополиплоидов) или экологических рас нельзя исключить возможности неоднократного параллельного возникновения на разных местах (политопное происхождение).
Рис. 14.17. Современное общее распространение семейства буковых (Fagaceae) с ареалами рода Fagus и родственного ему семейства Nothofagaceae с ареалами рода Nothofagus,а также ископаемые находки (+, х) обоих родов

14.2.1.3. Плотность заселения ареалов
Высокая плотность заселения указывает на то, что для данного вида область экологически оптимальна. При этом часто достигаются более сильная его экспансия на различные местообитания (максимальная экологическая амплитуда) и наибольшая генетическая диверсификация (многообразие форм, центр вариабельности). Аналогично и у родов или семейств, для которых регионы с особенно большим видовым богатством могут считаться центрами разнообразия. Это также области процветания и соответственно поддержания таких таксонов, но они не обязательно представляют собой центры происхождения. По всем этим критериям можно установить так называемое Ядро, или центр ареала.
Очанка крошечная (Euphrasia minima) произрастает в Центральных Альпах от горного до нивального пояса (1 400 — 3 100 м); ее максимальная встречаемость, наиболее широкая экологическая амплитуда и центр варьирования приходятся на нижний альпийский пояс между 2000 и 2 300 м (49 фенотипов по сравнению с двумя на верхней и восьмью на нижних границах распространения). Пример центра разнообразия рода — род Ononis, представленный наибольшим количеством видов в юго-западном Средиземноморье (рис. 14.18), для семейства — Rubiaceae (Мареновые), с высоким родовым разнообразием в теплых и влажных тропиках и прогрессирующим его убыванием при переходе к более сухим или холодным областям.
Рис. 14.18. Центр разнообразия и зоны снижения числа (обозначены цифрами) видов для рода Ononis (стальник, Fabaceae)

14.2.1.4. Географические связи между ареалами
По пространственному положению ареалов и их центрам можно объединить близкие виды в определенные (иерархически сгруппированные) геоэлементы. Центры распространения наиболее важных, участвующих в сложении флоры Центральной Европы геоэлементов представлены на рис. 14.19. Ареалы многочисленных видов, относящихся к определенным геоэлементам, суммируют, что дает возможность определить их центр распространения; это показано на рис. 14.20 на примере циркум-полярно-арктоальпийского геоэлемента во флоре Скандинавии.
Рис. 14.19. Флористические области западной части Евразии и Северной Африки в пределах Голарктического флористического царства Северного полушария: области; подобласти; иные линии границ (провинции и т. д.): циркумарктическая, циркумбореальная, центрально-европейская (с атлантической, субаглантической, среднеевропейской, альпийской, карпатской), понтийско-южносибирская (с паннонской, среднесибирской и т. д.), макаронезийско-средиземноморская (с субсредиземноморской, кавказской и т. д.), восточно-туранская (с аралокаспийской и т. д.), сахаро-синдская. N — северо; S — южно; О — восточно; W— западно; Z — центрально
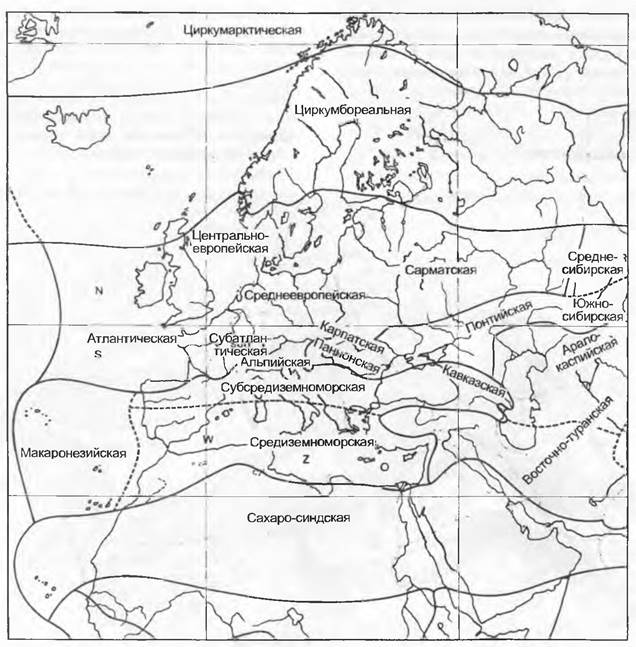
Род Fagus относится к голарктическому, вид Fagus sylvatica — к центрально-европейскому элементу, близкородственный Fagus orientalis имеет реликтовый ареал в горах Кавказа (примеч. ред.) и на Черном море. Род Laurus представлен в Европе двумя видами: L. nobilis и L. azorica (= L. canariensis); первый входит в средиземноморский геоэлемент, второй — в макаронезийский. Два близкородственных вида этого реликтового рода подчеркивают тесные связи между макаронезийской и средиземноморской флорами (единый ареал в третичном периоде). Охват всего тропического и, соответственно, подчиненного, более узкого неотропического элемента можно продемонстрировать на примере семейств Аrесасеае (пальмы) и Вrоmеliасеае.
Рис. 14.20. Распространение 50 циркумполярно-арктоальпийских скандинавских видов в Северном полушарии. Концентрация соответственно снижается к краям, а обеднение этого геоэлемента показано штриховкой различной плотности

14.2.1.5. Климатические флористические зоны
Широтные зоны. Многие ареалы можно опознать по тому, что они примерно приурочены к определенным широтам и имеют форму, более или менее приближающуюся к поясной (рис. 14.21, см. рис. 14.17). Поэтому надежной системой понятий для районирования и описания ареалов поясного типа, последовательно сменяющих друг друга от экватора к полюсу и соответствующих температурным градиентам, служат ботанико-географические (флористические) зоны (рис. 14.22). При этом если начать с тропической (t) и граничащей с ней субтропической (subtrop) зон, далее на север выделяют меридиональную (m), субмеридиональную (sm), умеренную (temp), бореальную (b) и арктическую (а) зоны, на юг — южную (austr) и антарктическую (antarkt) зоны, которые соответствуют зонам Северного полушария: первая — всем зонам от меридиональной до умеренной, вторая — бореальной и арктической.
Рис. 14.21. Формирование ареала и историческая дифференциация жизненных форм рода Carlina (колючник, Asteraceae). Канарские и соответственно восточно-средиземноморские таксоны подрода Carlowizia и подрода Lyrolepis U а, b, слева внизу) — реликтовые и относительно более древние (исходные), другие (2а, b и За, b, с) — циркумсредиземноморские и соответственно средиземноморско/монтанно-центрально-европейские, расширяющие ареал и прогрессивно производные. Аллопатрическая дифференциация С. vulgaris на западе и С. biebersteinii (лист изображен сверху справа) на востоке.

Континентальность — океаничность. Дальнейшей дифференциации можно достичь, если считать, что в океаническом секторе годовые и суточные значения температуры и влажности наиболее уравновешены, тогда ареалы по положению относительно него характеризуются как эуокеанические (euoz), океанические (oz), субокеанические (suboz), субконтинентальные (Subк), континентальные (к) и эуконтинентальные (еuк). Океанические ареалы располагаются вблизи побережий и находятся под сильным влиянием моря, гумидного климата с малыми годовыми амплитудами температур; континентальные apeaлы находятся в удаленных от побережья районах с сухим климатом и большими температурными амплитудами.
Рис. 14.22. Флористические зоны и секторы океаничности биосферы Земли (сокращения см. в тексте)

Высотные зоны. Принимая во внимание высотные пояса (см. рис. 14.44, раздел 15.1.2) для Центральной Европы: равнинный (pI), холмисто-равнинный (coll), монтанный (mо), субальпийский (salp) и альпийский (alp)], можно наконец получить трехмерное описание ареала, представить его в виде формулы и свести похожие ареалы в соответствующие типы.
Формула sm/mo-b(k) EURAS характеризовала бы ареал Pinus sylvestris как субмеридионально-монтанный до бореального, умеренно континентальный, евразиатский, область распространения Quercus robur можно описать формулой sm/mo- temp(suboz) EUR, т. е. как субмеридионально-монтанную до умеренной, слабо субокеаническую, европейскую, aFagus sylvatica— m/mo-temp(oz) EUR, т.е. как меридионально-монтанную до умеренной, океаническую, европейскую.
14.2.1.6. Спектры типов ареалов
Ареалы и типы ареалов (см. рис. 14.14) можно осмыслить и сравнить по очень различным критериям, теперь это лучше делать с помощью компьютеров. Если рассчитать доли известных типов ареалов в определенных флорах или единицах растительности, полученные спектры типов ареалов позволят сделать важные Выводы относительно их структуры и происхождения.
Высокая степень эндемизма на Гавайских островах (почти 20% родов и 90% видов наземной флоры сосудистых растений являются эндемиками) подчеркивает, к примеру, относительно древний геологический возраст этого архипелага, отсутствие древних связей с материками и значительную самостоятельность эволюции тамошней живой природы из ограниченного числа различных исходных особей, которым удалось проникнуть туда дальними путями расселения. Спектры типов ареалов смешанных лиственных лесов самой восточной части Европы показывают, что на Урале относительно высокая доля бореальных и южно-сибирских видов, напротив,
далее к югу, в Донецком бассейне — субсредиземноморского и понтийского геоэлементов. Эти различия будут понятны, если принять во внимание близость соответствующих рефугиумов ледникового времени, их роль в постгляциальном восстановлении лесов и современную климатическую ситуацию в данных районах.
14.2.2. Распространение
14.2.2.1. Возможности миграции
Растения, ведущие прикрепленный образ жизни, могут образовывать и расширять свои ареалы только в том случае, когда они образуют единицы расселения, так называемые диаспоры (например, Споры, семена, закрытые плоды, вегетативные единицы распространения — луковички и выводковые Почки и т. п.; см. 11.2, семена, плоды), с помощью которых они укореняются на новых местах. Подобные единицы расселения продуцируются часто в большом количестве и специализируются либо к автохории, либо к разносу ветром, водой или животными (см. 11.2, семенные растения). Естественным образом распространение успешно происходит только на довольно малые расстояния, однако иногда это приводит к дальнему распространению.
Расселение одного вида (миграция) может происходить в форме миграционного фронта, мелкими «шагами» или большими «прыжками», как вероятностный феномен с образованием форпостов (рис. 14.23).
Рис. 14.23. Миграция растений может осуществляться в форме широкого (А) либо узкого (В) фронта распространения (мелкими «шагами») или путем создания форпостов (С) даже из реликтовых местообитаний, а также большими «прыжками» (последнее — более частый случай, причем между ними существуют все переходы, и типизация зависит от градаций)

Мелкие или снабженные хорошими приспособлениями для переноса диаспоры могут отрываться, высоко подниматься штормовыми ветрами и переноситься ими на сотни километров. Перелетные и водоплавающие птицы при случае переносят диаспоры трансатлантическими путями, и ряд свежих дизъюнкций между Южной Америкой и Африкой часто возникал именно таким образом (например, эпифитных кактусов рода Rhipsalis с сочными плодами, которые даже достигли Мадагаскара и Шри-Ланки). Кокосовая пальма (Cocos nucífera) благодаря своим плавучим плодам, снабженным непроницаемым для волы твердым ядром и богатым жирами эндоспермом, долго не теряющим всхожести в соленой воде, смогла завоевать ареал, охватывающий все тропические побережья мира (см. 15.2.16).
Самый лучший модельный объект для оценки скорости и успеха расселения в далеко отстоящие биотопы — это островные местообитания как в море, так и на суше, где, к примеру, горные вершины, подобно островам, отстоят изолированно друг от друга на далекие расстояния. Особенно поучителен пример Кракатау. После мощного вулканического извержения, случившегося в 1883 г., на этой группе островов, расположенных между Явой и Суматрой, было уничтожено все живое. В 1934 г. там уже закрепился 271 вид наземных растений, переселившихся с территорий, находящихся на расстоянии 45 — 90 км.
В общих чертах цепочки островных местообитаний представляют собой миграционные коридоры (островные или горные «прыжки»). Так, индомалайская островная страна во многих случаях послужила путем распространения между Восточной Азией и австралийско-западно-тихоокеанскими территориями. Аналогично вдоль Кордильер на западе Северной и Южной Америки или по вулканическим плато Восточной Африки происходил длительный флористический обмен между Северным и Южным полушариями, один из самых недавних в прошлой истории Земли (например, род Alnus, который успел дойти через Центральную Америку до Южных Анд).
За прошедшие столетия естественные механизмы дальнего распространения видов постепенно были доведены до предела деятельностью человека. В настоящее время вид может «передать» свои диаспоры на любом авиарейсе или в багаже туриста, и все страны света для него уже открыты. Так, естественные леса из Metrosideros на Гавайях через несколько десятилетий могут пасть жертвой инвазии Myrica flava с Канарских островов, вида, который высадили там в 1970 г. и который с тех пор был полностью без контроля. Род Eucalyptusсейчас распространен во всех теплых странах света.
Однако транспортный потенциал диаспор надежно определяется только одним из многих факторов, а именно величиной ареала и его состоянием. В пределах многих богатых родов существуют бок о бок узколокализованные виды, или дизъюнктивные палеоэндемики, и очень экспансивные сорные растения, и те, и другие имеют очень сходные диаспоры (например, в родах Taraxacum). Многие ГРИБЫ, мхи и папоротники со своими пылевидными спорами имеют сходные по границам и часто дизъюнктивные ареалы, такие же, как и у семенных растений с тяжелыми диаспорами (например, папоротник олений рог, Asplénium scolopendrium, и виды рода Tagus, см. рис. 14.17). Необходимо привести факторы, которые снижают фактическую миграцию по сравнению с потенциальной.
14.2.2.2. Препятствия для миграций
Прежде всего учтем быстрые изменения ареалов, связанные с резкими изменениями во внешней среде, например, в периоды начала и окончания оледенений или в случае антропогенно обусловленного потепления климата. Среди многих причин, которые тем не менее тормозят миграцию видов и сообществ при устранении климатических границ и физических барьеров для распространения, особенно важны три.
• Распространяется не «растительность», а отдельные виды, которые должны потом утвердиться в совершенно новой для них биотической среде. Только в крайне невероятном случае, если все виды фитоценоза быстро и одновременно переселятся или будут так тесно сопряжены друг с другом, что могут мигрировать только совместно, фитоценоз «передвинулся» бы целиком.
• При миграции необходим партнер. У двудомных, или самостерильных, видов для продолжения рода необходимо по меньшей мере два разных экземпляра растения. Именно поэтому на океанических островах самоопыляющиеся покрытосеменные представлены сверх нормы, ведь даже из одного растения может сформироваться популяция (см. 10.1.3). Часто (например, на Гавайях) наблюдается вторичная двудомность как выход из возможного близкородственного скрещивания (инцухта). Но даже при преобладании автогамных видов наличие большего числа различных исходных особей одного вида благоприятно сказывается на поддержании их в хорошей форме. К тому же более 90% всех видов растений образуют микоризу, и совершенно понятно, что эти связи гриба и растения высоко специализированы и совместимость с партнером генетически очень узко ограничена. Многим видам растений необходимы специальные опылители или даже патогенные организмы, которые должны подавлять возможных конкурентов и т. д.
• Почвы и весь остальной экотоп не мигрируют. Почвы формируются под воздействием исходной материнской породы, климата и растений. Таким образом, они как бы являются реакцией на многолетнее существование определенного растительного сообщества, которое изменяется вместе с почвой (сукцессия; см. рис. 14.1, 14.33, раздел 14.3.2), что имеет значение для микросреды обитания с климатической точки зрения.
Даже при «открытых каналах миграции» и соответствии климата и субстрата новых местообитаний перечисленные обстоятельства приводят к тому, что скорость миграции существенно медленнее, чем можно было бы ожидать от распространения диаспор. При сохраняющихся условиях среды миграция лимитируется количеством диаспор только в самых редких случаях. Стойкая натурализация нового вида растений сталкивается всегда с гораздо большими трудностями, чем транспорт диаспор. Не
скольким тысячам заносимым случайно в Европу видам (неофитам) противостоят только несколько сотен действительно натурализовавшихся иноземных видов покрытосеменных. В новых условиях они никогда не встречают (или очень долго не находят) растительные сообщества, сходные с теми, из каких они пришли путем миграции. Миграция приводит, как правило, к созданию новых растительных сообществ, состоящих из смеси элементов, относящихся к разным этапам времени внедрения и происходящих из разных областей.
И горечавка бесстебельная (Gentiana acaulis S.I.), и эдельвейс (Leontopodium alpinum) относятся к элементам современной центрально-европейской горной флоры. Но в то же время, если Gentiana acaulis s.l. на этой территории является коренным (автохтонным) видом и принадлежит к позднетретичному ядру альпийской флоры, то Leontopodium alpinum в Альпах относится к мигрантам ледникового периода, он происходит из Центральной Азии и при отступании ледника мигрировал в Высокие Альпы из холодно-сухих в ледниковое время предгорий.
Требования к хорошему расселению семян, с одной стороны, и к удовлетворительному снабжению проростков, с другой, находятся в противоречии между собой (см. 14.1.3). У пионерных растений много мелких семян, но они всходят только на свету и долго не сохраняются (у деревьев, например, ивы или сосны), у видов поздних сукцессионных стадий преобладают семена часто не столь многочисленные, но крупные и хорошо снабженные резервными питательными веществами, прорастающие в тени (т. е. в условиях конкуренции) и длительно сохраняющие всхожесть (например, у бука или дуба). Все эти определяющие успех миграции факторы отбирают виды широкого профиля, элементы первичной сукцессии, в связи с чем мир переполнен рудеральными «агрессорами», которые только изредка (но, к сожалению, это все же случается) вторгаются в местную растительность, если она достаточно «открыта». Примеры такого внедрения — суккуленты Нового Света (Agave, Opuntia) в Средиземноморской области или «оккупация» полынных степей Большого Бассейна запада Северной Америки евразиатским злаком Agropyron desertorum, из-за
которого там повышается частота пожаров, изменяющих всю систему (см. рис. 13.26).
Наконец, организмы с быстрой сменой поколений (например, Бактерии или короткоживущие терофиты) скорее способны к быстрому расселению, чем те, у которых до стадии плодоношения должны пройти долгие годы (например, у долгоживущих древесных растений). Скорость перемещения вида зависит также от биологии его размножения и длительности существования поколений, от биотических взаимодействий и действительных условий нового жизненного пространства.
Постгляциальная миграция Fagus sylvatica (бука) из окраин Альп до Балтийского и Северного морей (700 км) заняла почти 3 000 лет, для завоевания им здесь доминирующего положения среди других лиственных пород понадобилось еще 2 000 лет (11.3.2.4); в западной Скандинавии расселение бука к северу еще не завершилось. Исключительно путем вегетативного размножения элодея канадская, или водяная чума (Elodea canadensis), появившаяся в Ирландии в 1836 г., а в Берлине в 1859 г., взрывными темпами размножилась в европейских водах и расселилась за счет коренных водных растений, хотя в последние десятилетия заметно ее явное отступание (причина: нематода, поражающая конус нарастания растения).
14.2.2.3. Распространение и модификация
Вместе с пространственным распространением видов или их групп часто происходят увеличение разнообразия и прогрессирование признаков, которые следуют повторяющимся типам (см. 10.3). Три из них особенно типичны.
✵ Многие таксоны в исходных районах произрастания являются деревянистыми растениями, в то время как на осваиваемых ими территориях интенсивная диверсификация порождает травянистые таксоны. К этому вынуждает то. что на пути миграции виды с коротким жизненным циклом имеют более высокую вероятность достичь отдаленных территорий.
Пример — эволюция тропических преимущественно древесных Caesalpiniaceae в сторону более прогрессивных травянистых Fabaceae умеренного пояса. У чертополохов рода Carlina (Asteraceae, см. рис. 14.21) можно увидеть следующие линии эволюции: от вечнозеленых канделябровидных кустарников с мелкими корзинками, распространенных в поясе лавролистных лесов макаронезийских островов (например, С. salicifolia, subg. Carlowizia. I а) и многолетних крупных трав с сильно одревесневшими основаниями побегов, растущих в Средиземноморье (например, С. corymbose agg., 2а). далее или в сторону средиземноморских однолетников (например, С. racemosa, 2и), или к малолетним и только раз в жизни цветущим розеточным растениям (С. vulgaris agg., Зе), или, наконец, к совершенно лишенным стебля травам с крупными корзинками (С. acaulis, За; С. acanthifolia, 3b), достигшим при расселении умеренных областей Западной Евразии.
✵ Очень часто распространение близкородственных групп сопровождается скачкообразными изменениями хромосомных чисел, закрепляющимися в потомстве.
Примеры — дисплоидные ряды (см. 10.1.2.2) у Myosotis, Chaenactis, Haphpappus или развитие от диплоидов к полиплоидам, как у Asplénium, Biscutella, Galium, Achillea, Aegilops (см. 10.3.3.4).
✵ Сходным образом протекает и прогрессивное изменение биологии размножения, почти всегда в определенном направлении (см. 10.1.3.3): от аллогамии к автогамии, от полового размножения к апомиксису, или же происходят интерпретируемые с позиций кладогенеза изменения в последовательности ДНК.
14.2.2.4. Сокращение ареалов
За фазами прогрессирующего расселения видов растений часто следуют фазы приостановки данного процесса или даже регресса, что приводит к сокращению ареалов. Этого нельзя увидеть только при рассмотрении современных ареалов, но подтверждается и даже датируется на основе ископаемых находок. Многие виды, которые были оттеснены на территории рефугиумов во время климатических колебаний в конце палеогеннеогенового периода или во время плейстоценовых оледенений, так и не восстановили свои первоначальные ареалы, несмотря на улучшение климата, или восстановили их только частично. Нередко реакцией на это была потеря внутривидового генетического разнообразия и, таким образом, способностей к приспособлению. Из третичной лесной флоры (см. рис. 1 1.298), например, в Европе сохранились только остаточные ареалы Picea omorica или Aesculus hippocastanum на Балканах. Полностью вымерли в Европе роды Ginkgo и Magnolia. Первый выжил лишь в Юго-Восточной Азии, второй — на востоке Северной Америки и в Восточной Азии (откуда снова был интродуцирован в европейские сады; см. рис. 14.16, 14.24).
Рис. 14.24. Распространение семейства Magnoliaceae — современное (показано штриховкой) и в прошлые периоды истории Земли (• — ископаемые находки за пределами современного ареала, с верхнего мела, в палеоген-неогеновом периоде и до плейстоцена)

Сжатие и сокращение ареала рода Ginkgo — особенно выразительный пример подобной динамики ареалов в геологических масштабах (см. рис 14.16). В верхней юре произрастание Ginkgo было ограничено Центральной Азией. Начиная с верхней юры до раннего палеогеннеогенового периода род достиг Шпицбергена и Аляски и был представлен большим числом форм. Его вымирание началось в конце палеоген-неогенового периода на территории Северной Америки, а затем и в Европе и привело к современному реликтовому ареалу только одного выжившего вида — Ginkgo biloba, «живого» ископаемого, произрастающего естественно только в нескольких местах в Китае. По ископаемым находкам твердо установлено также отступание арктоальпийских растений в горы и на север Европы со времени последнего оледенения (например, Salix herbácea). В последние десятилетия многие растения верховых болот (например, Ledum или Scheuchzeria) или сухих малопродуктивных лугов (например, некоторые орхидеи и виды рода Pulsatilla) из-за прогрессирующего обводнения или, наоборот, иссушения значительно отступают. Последняя популяция Marsilea quadrifolia (см. рис. 11.160) в Германии была уничтожена недавно, в 80-е годы XX в., из-за сооружения мусорной свалки.
14.2.3. Причины, обусловливающие границы и площадь ареалов
Границы ареалов обусловлены не только историческими и физическими причинами (например, взаимного размещения материков и морей), но и многими экологическими, поскольку связаны с условиями современного климата и почвенным покровом (см. рис. 12.5, 14.1), а также с экофизиологическими свойствами видов
растений. Значительная часть ареалов обусловлена связанными с температурой зонами растительности или придерживается секторов, обусловленных океаничностью климата (см. 12.5.2.2). Многократно проводились попытки наложить границы ареалов на определенные изолинии климатических показателей, чтобы рассматривать последние как климатические факторы формирования границ (рис. 14.25). Ввиду комплексной природы климатических и почвенных условий, а также сочетания их с явлениями конкуренции подобные попытки тем не менее остаются проблематичными.
Имеющие ступенчатую форму восточные границы распространения Fagus sylvatica (бука) и Quercus robur (дуба черешчатого) явно обусловлены климатически (см. рис. 11.259). Дуб значительно устойчивее к экстремальным температурам и засухам, чем бук. У атлантического падуба (Ilex aquifolium), имеющего близкородственные виды не только в западной, но и в восточной Евразии и в Гималаях (см. рис. 14.25), южная и восточная границы совершенно явно обусловлены возрастанием летней сухости. Суровость зим (континентальность!) также связана, но не напрямую, с указанными температурными кривыми. Решающими здесь очевидно являются низкие зимние температуры (менее -15 °С), вероятность произрастания коррелирует с этой изолинией, ощутимые повреждения морозами влекут за собой тяжелые последствия и обусловливают ослабление конкурентоспособности. Ареал Hex заметно сузился после экстремальной зимы 1928/29 гг.
Рис. 14.25. Распространение падуба (Ilex aquifolium) и близкородственных видов рода в западной части Евразии. Для сравнения приведены январская изотерма 0 °С по Н. Meusel et al. и изотерма по Климатическому атласу Европы (Юнеско, 1970)

Даже когда ареал вида из чисто прагматических соображений называют замкнутым, вид в его пределах всегда ограничен своими специфическими местообитаниями, причем размеры и частота встречаемости этих местообитаний определяют внутреннюю плотность заселения ареала. Значительное перекрытие на плоской проекции ареалов разных таксонов вовсе не означает, что эти виды или группы видов имеют одинаковые требования к условиям среды. Ареал всегда неизбежно покрывает весь спектр типов местообитаний ландшафта, различных по положению в рельефе, экспозиции, влажности и субстрату.
У бука, дуба и сосны в Центральной Европе ареалы значительно перекрываются. Однако в пределах одной области они заселяют тем не менее совершенно разные местообитания, поэтому часто не растут в одних и тех же сообществах, но являются характерными видами буковых, дубовых и сосновых лесов. Если сравнить определенные в чистых культурах примерные оптимальные значения влажности и pH почвы, то между этими тремя видами обнаружится очень мало различий. В смешанной же культуре светолюбивый дуб отступает от теневыносливого бука в краевую зону благоприятных для него по условиям мест, а еще более светолюбивая сосна почти полностью подавляется. Сосна ни в коем случае не «предпочитает» в Центральной Европе более сухие места, или сырые болотные, или участки с бедными и кислыми почвами, но она может выдерживать такие экстремальные условия, потому что там отсутствует конкуренция со стороны более требовательных к условиям лиственных пород (см. 12.1).
Некоторые виды отличаются удивительно широкой амплитудой по отношению к климату и в пределах ареала находятся в самых различных климатических зонах, что часто наблюдается вместе с явственной экотопической дифференциацией популяций. Хороший пример из этой области имеется также для лесообразующих пород: ареал сосны простирается от Южной Испании до Лапландии, бука — от Сицилии до Северной Скандинавии (см. рис. 11.259). Это стало возможным, с одной стороны, благодаря экологической компенсации (на юге они занимают северные склоны вместо южных или поселяются на больших высотах), с другой — образованию различающихся экотипов (см. рис. 10.27, В). Fagus sylvatica в центральной части своего ареала распространен от пояса холмистых равнин до верхнего горного, но на юге он ограничивает свое распространение более холодными горными лесами, а на северной границе растет на почвах низменных равнин. Горные растения, например, осока твердая (Сагех firma) или растущая в поясе криволесья горная сосна (Pinas mugó), могут часто селиться в условиях холодно-влажного местного климата даже на равнинных поверхностях.
Часто на основании локального распространения вида и его связи с местообитанием можно судить о его общем распространении и наоборот; то и другое, являясь как бы «общим знаменателем», лежит в основе экофизиологической шкалы потребностей и допустимых пределов существования вида или группы видов (часто описывается как норма реакции). А потому карты распространения можно было бы употребить при соответствующей критической оценке как показатель определенного фактора местообитания. В какой мере они отражают причинные связи, вопрос масштаба. Мелкий масштаб дает Общее представление о требованиях к макроклимату. Более крупный масштаб с большей вероятностью отражает эдафические причины, обусловливающие распространение. Связь между ареалом и современными условиями окружающей среды особенно бросается в глаза на примере видов, жестко привязанных к определенному типу почвы. С давних пор известны растения засоленных, песчаных, известковых и щебнистых грунтов 4см. 6.2.2.4, 13.6.6).
Как пример, сошлемся здесь на солевыносливые (например, Salicornia europaea agg., Glaux maritima и Aster tripolium) и предпочитающие пески (например, Salsola kali и булавоносец, Corynephorus canescens) растения морских побережий и удаленных от моря участков. Только на серпентиновых скалах произрастают некоторые папоротники, например, Asplénium adulterinum. В Альпах встречаются лишь на известняковых или соответственно на силикатных породах близкие, но не близкородственные (псевдовикарирующие, см. рис. 14.14, Е) пары видов Rhododendron hirsutum и R. ferrugineum, Gentiana clusii и G. acaulis s. str. (= G. kochiana). При этом решающую роль играет реакция почвы (см. 6.2.3). Среди полевых сорняков дикая редька, Raphanus raphanistrum, растет на более или менее кислых почвах, а полевая горчица, Sinapis arvensis, напротив, на основных или очень слабокислых. Постоянство по отношению к почвам вообще может меняться со сменой климатических условий: многие виды, которые в более сухих и теплых районах растут на различных почвах, в районах с холодным и влажным климатом становятся кальцефилами, т. е. здесь та же основная или нейтральная реакция почвы имеется только на известняках.
14.2.4. Биоразнообразие и стабильность экосистем
Изменчивость и многообразие организмов — причина и результат эволюции, основное свойство всего живого. Темпы современного, антропогенно обусловленного снижения биологического разнообразия эквивалентны масштабному вымиранию (great extinction) видов, имевшему место в истории Земли предположительно по причине столкновения с метеоритом. Благодаря конвенции, принятой в 1992 г. в Рио- де-Жанейро, понятие биоразнообразия и нависшей над ним угрозы выдвинулось в сферу общественных интересов.
Существует много мотивов для поддержания биологического разнообразия, каждый из которых имеет достаточную внутреннюю ценность: этика (защита жизни, как таковой), экологическое значение (см. ниже), экономическое значение (Питание, безопасность, чистая питьевая Вода, природные Материалы), культурное наследие (старые, созданные человеком экосистемы), эстетическая ценность (красота природы) и др. В этом разделе изложены некоторые причины, основанные на биологических и экологических мотивах. Прежде всего необходимо оговориться, что биология в данном случае не имеет первенства, сохранение биоразнообразия не нуждается в оправданиях с естественно-научных позиций, но это тем не менее может дать ценные дополнительные аргументы, хотя фактическая ситуация местами еще очень неудовлетворительная.
14.2.4.1. Биоразнообразие
Биологическое разнообразие, кратко называемое биоразнообразием, охватывает все многообразие «биологических единиц» в определенный промежуток времени в пределах заданного пространства. «Биологические единицы» могут быть представлены генетически различающимися индивидумами некоей популяции, а также таксономическими единицами (вид, род, семейство), жизненными формами (см. 14.3.1, рис. 14.19) и функциональными типами (см. 12.5.1.3), сообществами и экосистемами как зеркальным отражением многообразия биотопов в ландшафте (см. рис. 12.8, 12.9, 12.13). Здесь эта тема изложена только примерно. В основном рассмотрены в первую очередь ботанические таксоны (как правило, виды), не умоляя, однако, значения разнообразия биологических единиц других уровней и категорий. Важность внутривидового разнообразия обсуждалась в предшествующих разделах (см. 10.1, 12.4).
Общее количество видов покрытосеменных (таксономическое разнообразие) сейчас оценивается в 240 000, мхов — почти в 24 000, папоротников — в 10 000 и голосеменных — около 800. На протяжении всей истории Земли биоразнообразие возрастало.
Разнообразие видов внутри одного растительного сообщества называют α-разнообразием, изменение комбинаций видов между разными сообществами одной местности — β-разнообразием (разнообразие сообществ видов). Даже в пределах относительно однородного по условиям внешней среды местообитания может быть дифференциация совокупностей видов вследствие только того, что виды имеют тенденцию к небольшому радиусу распространения диаспор, встречаясь как бы «сгустками», что в большинстве случаев приводит к небольшому a-разнообразию и высокому β-разнообразию. На площади 100 м2 могут совместно произрастать в зависимости от морфотипов (размеров растений) до 200 видов (травянистых). На близком к естественному сенокосном лугу для такой площади типичное число видов — около 30, на очень богатых видами известняковых лужайках— 80 — 100. Водном из самых разнообразных по видовому составу лесов Земли, Пасо в Малайзии, на площади 2 га произрастает 276 древесных видов (только со стволами толще 10 см, а всего на этой площади растет 1 169 видов, по Т. Kira; сюда еще не включены виды кустарников, крупных трав, эпифитов и большинства лиан).
Биологическое разнообразие распределено по поверхности Земли неравномерно. Число видов, или таксономическое разнообразие, относительно больших (> 1 км2) единиц площади можно рассматривать глобально следующим образом:
✵ от полюса к экватору;
✵ от территорий с неблагоприятным (чрезмерно холодным или сухим) климатом к районам с благоприятным (относительно теплым и влажным) климатом;
✵ от нестабильных периодов в истории Земли (например, оледенений) к стабильным;
✵ от однообразных территорий к сильно дифференцированным, с высоким разнообразием биотопов.
Это хорошо видно при взгляде на мировую карту биоразнообразия (число видов сосудистых растений на 10000 км2, см. форзац). Одной из причин того, почему на этой схеме центрами биоразнообразия оказались области с высокими горами, является масштаб карты, где-за единицу принят квадрат размером 100 x 100 км, при этом в один квадрат сетки умещается вся горная система. В тропических горах это имеет большое значение, поскольку в один квадрат сетки включается биоразнообразие и прилежащих тропических равнин, и всех умеренных поясов вплоть до границы снегов. Из этого понятно, какое значение имеет высокое разнообразие биотопов для общего регионального биоразнообразия. Более нигде не имеется на относительно небольшой площади столь высокого разнообразия организмов, и при решении природоохранных задач следует учесть в первую очередь горы, особенно тропические.
Эта схематическая карта с 10-балльной цветовой шкалой, дифференцирующей зоны наземного биоразнообразия, основана на первичной и довольно грубой экстраполяции доступных в настоящее время, различающихся по достоверности и еще далеко неполных представлений о флоре земного шара. Бросается в глаза 6 глобальных центров разнообразия сосудистых растений, все они лежат в тропических и субтропических горных районах: (1) Чоко — Коста-Рика; (2) тропическая часть восточных Анд; (3) приатлантическая Бразилия; (4) восточные Гималаи — Юньнань; (5) север Борнео; (6) Новая Гвинея. На настоящий момент особенно разнообразными считаются флоры Венесуэлы и Новой Гвинеи (20 000 — 30 000 видов сосудистых растений).
Общие причины снижения биоразнообразия при удалении от экваториально-тропической зоны и с увеличением высоты в горах (в Альпах и горах Скандинавии выше верхней границы леса богатство цветковых растений падает почти на 40 видов через каждые 100 м подъема) следующие: (1) уменьшение доступной поверхности (чисто геометрически, поскольку Земля представляет собой шар, а гора — конус); (2) снижение длительности вегетационного периода (52 недели в гумидных тропиках, 10 недель выше границы леса в горах или в арктических тундрах); (3) увеличивающееся воздействие морозов (селективный отбор); (4) увеличение ярко выраженного действия масштабных климатических колебаний (оледенения). Если принимать во внимание пространственные и временные ограничения эволюционных процессов (1 + 2), то едва ли видовое богатство некоторых горных флор уступит равнинным.
Классическое объяснение того факта, почему вообще многие виды могут произрастать совместно на узком пространстве, содержится в принципе конкурентного исключения (competitive exclusion), или в принципе Гаузе: видам трудно существовать совместно, если они имеют сходные жизненные формы и сходные потребности в ресурсах (см. Приложение к гл. 12). Структурная и функциональная дифференциация, напротив, позволяет использовать дополняющие друг друга ресурсы (дифференциация ниш). Из этого можно сделать вывод, что чем больше видов может существовать совместно, тем уже их ниши (см. 14.1.2). Далее предположим, что разнообразию способствует влияние серьезных нарушений, особенно нагрузки со стороны травоядных и патогенных организмов на виды, которые без подобных нарушений стали бы доминирующими. Значительный дефицит питательных веществ также способствует увеличению разнообразия, поскольку быстрорастущие, требовательные к питанию виды при этом не могут доминировать и становится возможным более богатое сочетание видов. В реальности на все эти аргументы находятся и противоположные примеры. Особенно производит впечатление довольно хорошо обоснованный тезис, что, противореча принципу Гаузе, как раз широкие ниши тропических деревьев (перекрывающиеся ниши) делают возможным сосуществование очень многих видов в девственных тропических лесах.
Наряду с просто числом видов S применяют ряд математических индексов разнообразия, из которых наиболее употребительным является индекс Симпсона D. Он выражается как D ![]() где х1 — доля площади (покрытие) i-гo вида.
где х1 — доля площади (покрытие) i-гo вида.
D будет равно S в случае, если все виды занимают одинаковую площадь. D обычно значительно меньше S, когда в растительности доминирует небольшое число видов. Таким образом, D выражает и асимметрию в обилии видов. Виды, которые хотя и имеются в наличии, но почти не вносят вклада в общее покрытие (биомассу, экосистемные Функции), рассматриваются также в качестве вносящих минимальный вклад в биоразнообразие, что не должно соответствовать их «весу» в природоохранных задачах.
14.2.4.2. Биоразнообразие и функционирование экосистем
Попытка количественно оценить значение числа видов (разнообразия наиболее часто изучаемых биологических объектов) для процессов, происходящих в экосистемах, на первых порах привела к жестким противоречиям, конечная причина которых состояла в том, что не было достигнуто соглашения о едином временном масштабе. Говоря иначе, это означает, что взаимодействие биоразнообразия и экосистемных процессов зависит от масштаба. Для простоты вполне можно считать, что продуктивность (поскольку она несет интегрирующую функцию) замещает все прочие процессы.
Для начала приведем явно противоречивые факты: на однородно занятом экспериментальном поле продуктивность в среднем увеличивалась вместе с числом видов, одновременно выращиваемых в смешанном посеве. Этот эффект был наибольшим в посевах 1 — 4 видов, но с дальнейшим увеличением числа видов сходил на нет (A. Hektor и др. D.Tilman и др.). С другой стороны, влажно-тропические леса обнаруживают самое высокое видовое богатство, однако продуктивность в них не выше, чем в более бедных видами лесах умеренного или бореального пояса, если пересчитывать ее на фактически имеющееся в распоряжении время — на год (М. Huston; см. рис. 13.39). То же отмечается и в близких к естественным, малопродуктивных экосистемах дерновинных лужаек на бедных почвах в Европе с богатым видовым составом (Р. Грайм). На самом деле между этими экспериментами и полевыми наблюдениями совсем нет противоречий. Эксперименты показывают, что при обычных идентичных условиях (почвах) и в однородном насаждении большее число видов (в узком диапазоне разнообразия) имеет возможность произвести большую продукцию биомассы. Здесь просматривается эффект числа видов при заданном уровне ресурсов В других случаях обнаруженное (!) разнообразие представляет собой долгосрочную реакцию природы на заданный уровень ресурсов (и динамику нарушений). Эксперименты показывают, что потеря видов может иметь функциональные последствия. Согласно полевым наблюдениям, разнообразие видов саморегулируется в зависимости от ресурсов и нарушений.
При глобальном рассмотрении шансы на более высокое биологическое разнообразие имеют ландшафты с большим разнообразием биотопов, малыми экстремальными проявлениями климата и большим геологическим возрастом. Это, однако ничего не говорит о связи между биоразнообразием и функциями экосистемы конкретной парцеллы в пределах определенной зоны биосферы. Продуктивность измерить достаточно легко, но экологически она имеет меньшее функциональное значение, чем стабильность. Продуктивность может совместно с признаками климата и почвы способствовать стабильности, но так бывает не всегда. Стабильность (в смысле устойчивости, малой изменчивости) вообще не легко определяемое понятие, так как растительность не есть нечто статистически фиксированное, но подвержена длительным изменениям (например, сукцессионным циклам).
Надежный и более понятный критерий — обеспечение целостности экосистемы. Он ориентируется на долгосрочное сохранение почв с их влагоемкостью и запасанием минеральных веществ в экосистеме, а также на сохранение возможностей для будущего роста растений, принцип постоянства. Действительно ли высокое биоразнообразие обеспечивает целостность экосистемы лучше, чем низкое, в большой степени зависит от функциональных свойств, представленных в ней таксонов. Для этого рассмотрим несколько модельных представлений.
✵ Функционально различающиеся объединения таксонов равномернее и объемнее используют ресурсы (комплементарность ниш, англ. niche complementarity). Вследствие этого (при отсутствии нарушений) возникает сообщество видов с большей биомассой и другими преимуществами, например, со сплетающимися корнями, что позволяет более интенсивно и равномерно осваивать почву и при этом лучше защищать ее от эрозии.
✵ Благодаря разнообразию функционально различных типов растений, а также представленности последних многими видами, т. е. функциональной насыщенности, можно компенсировать последствия нарушений таким образом, что остаются гарантированными и защита почвы, и сохранение фонда минеральных веществ даже при выпадении из сообщества нескольких видов (страховая гипотеза).
✵ Частная форма страховой гипотезы — гипотеза «заклепок» (англ. rivet — заклепка). Она основана на том, что многие виды в экосистеме соединены крепко, как заклепки в самолете. Чем больше «заклепок», тем больше их может «выпасть» без ущерба для общей целостности системы. Особенность этой гипотезы состоит в том, что до момента достижения критического числа потерянных «заклепок», ведущего к катастрофе, потеря функциональности не заметна.
✵ Система с высоким разнообразием обладает повышенной способностью к экосистемной саморегуляции. Относительное противостояние стабильности экосистем изменениям условий внешней среды и биотической (или антропогенной) нагрузке основано, согласно этой гипотезе, на многообразных биотических взаимодействиях (интерференциях, обратных связях), которые связывают между собой всех членов экосистемы (например, питание консументов). На основе обратных связей обоюдно подавляются колебания численности популяций продуцентов и консументов первого и второго порядков.
Для этих убедительных и широко применяемых в компьютерном моделировании (по Д. Тильману, Е. Одуму) положений отсутствуют, однако, прямые доказательства из области естественных экосистем. Модель комплементарное ниш базировалась на искусственно созданных модельных системах травянистых растений. Наблюдались два разных эффекта: (I) посевы с более высоким видовым богатством (более четырех видов) с более высокой вероятностью и быстрее достигают высокого проективного покрытия (англ, cover), и, соответственно, их почвозащитная роль выше; (2) при сравнении только таких посевов с различным числом видов, которые достигают полного проективного покрытия, кроме того, при хорошем плодородии почвы с увеличением числа видов увеличивается и максимум сезонной биомассы (1-, 2-, 4-, 8-, 32-видовые смеси сравнивались в панъевропейском опыте, по A. Hektor и др., рис. 14.26, С). Сомкнутое луговое сообщество с более высоким числом видов и разнообразными спектрами типов побегов и листорасположения использует солнечный свет бесспорно лучше, чем монокультура или сообщество с малым числом видов. Точно так же многовидовое сообщество усваивает больше минеральных веществ из почвы, чем сообщество с низким разнообразием.
Рис. 14.26. Между биоразнообразием и функциями экосистемы может существовать связь. Это можно продемонстрировать на примере зависимой от биоразнообразия продуктивности (биомассы). Пять диаграмм символизируют пять возможных типов соотношений. Соотношения В — D действительны только для однородных формаций (дерновинные лужайки, лес); А — связи нет; В — линейная связь; С — связь в форме кривой насыщения, D — кривая с оптимумом; Е — пример, в котором происходит либо присоединение, либо выпадение нескольких видов («решающие» виды, например, бобовые или один вид дерева на лугу), либо изменение от более поздней сукцессионной фазы к более ранней (обычно после урагана или пожара), при этом функция изменяется скачкообразно. Примера из природы не существует только для варианта В
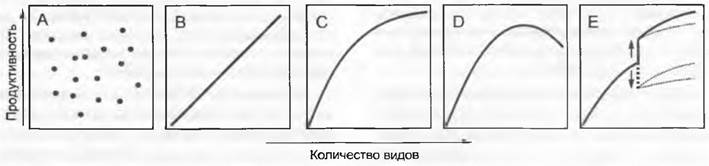
Генетически узкому (охватываемому только немногими видами) «занятию рабочей площади в экосистеме» может недоставать «страховой» насыщенности. Ее наличие — признак высокой конкурентоспособности, которая, как правило, тратится на устойчивость к стрессам и нарушениям. Но и очень простые, специализированные (устойчивые к стрессам и нарушениям) видовые сообщества могут гарантировать экосистемную стабильность (в смысле целостности) — это бореальные леса, некоторые травяные системы холодных регионов. К механическим воздействиям (например, ураганы, перевыпас) или пожарам часто они даже менее чувствительны, чем высокосложные сообщества видов, в которых после таких происшествий остаются свободные пространства и которые при подобных нарушениях бывают отброшенными в состояние более ранних стадий сукцессий.
Наконец, целостность экосистемы часто связана с наличием ключевых видов (key-stone species), т. е. когда выпадение одного-единственного вида (например, доминирующего вида древесного яруса) приводит к значительным изменениям. Итак, вопрос не только в том, как много видов могут (должны) выпасть (гипотеза «заклепок»), чтобы система потеряла равновесие, но и в том, какие это виды. Идентификация видов приобретает очень большое значение. На вышеупомянутые связи между биоразнообразием и продуктивностью (рис. 14.26) большое влияние оказывает отсутствие или наличие бобовых. Само по себе число видов, таким образом, недостаточный критерий для поддержания целостности и функциональности системы.
Основанное в большей степени на теоретической базе, нежели на фактах, представление о кибернетической «настройке» стабильности при высоком разнообразии путем саморегуляции находится в противоречии с особенной чувствительностью таких комплексных сложных экосистем, как, например, тропические саванны, в которых возрастание численности некоторых видов или их удаление (например, слонов или наземных хищников) приводит всю систему в состояние сукцессии (точно так же стимулирование или редукция циклических пожаров, которые связаны с образованием отмершей ветоши трав; см. 15.2.5).
Высокая, приобретенная за длительное время экосистемная стабильность, которая имеется в «Сердце» тропиков, часто обусловливает и высокое биоразнообразие, но последнее совсем не обязательно способствует высокой стабильности и целостности. Довод также необратимый. Для изначально мало разнообразных и регулярно нарушаемых систем «страховая» гипотеза имеет особенно большое значение для защиты почв и вместе с тем для обеспечения основ существования будущих поколений организмов. Невосполнимое выпадение одного вида может внезапно изменить такую систему и привести ее к ресурсному истощению (истощению почвы).
14.2.5. ФЛОРИСТИЧЕСКИЕ ОБЛАСТИ И флористические царства
Изучив размещение границ ареалов большого числа таксонов, нужно признать, что они распределены неравномерно, но в некоторых местах показывают довольно явное сходство. Этому соответствует тот факт, что между областями с гомогенной флорой и характерным видовым составом (А или В) лежат пограничные области с сильным флористическим градиентом и гетерогенным видовым составом (A/В). В большинстве случаев такие пограничные области совпадают с действительными границами распространения или с зонами, которые определяются изменениями климата. Две флористические области можно сравнить в отношении общих или различных видов и эндемичных для каждой области таксонов и количественно определить разницу как флористический контраст. Так, на флористической и ареалогической основе стало возможно провести пространственное расчленение всей биосферы (см форзац). Самые крупные объединения флористических областей — флористические царства. Различают шесть флористических царств мировой флоры с характерными для них семействами и наиболее важными родами.
• Голарктическое — самое большое флористическое царство, охватывающее все Северное полушарие Земли с арктической, бореальной, умеренной, субмеридиональной и меридиональной флористическими зонами: Pinaceae, Betulaceae, Fagaceae, Salicaceae, a также многочисленные Ranunculaceae и Rosaceae.
• Неотропическое — царство, занимающее субтропики и тропики Америки: Вгоmeliaceae (Tillandsia), Cactaceae; центр разнообразия Solanaceae (Solanum).
• Палеотропическое — царство, занимающее тропики и субтропики Африки и Азии вместе с Индомалайзией: Dipterocarрасеае (Юго-Восточная Азия), Combretaceae (Африка), Pandanaceae (пандановые), Zingiberaceae (имбирные) и центр разнообразия Moraceae (Ficus, Индомалайзия) и суккулентных Euphorbiaceae (Африка, Индия).
• Капское — маленькое, но очень характерное флористическое царство на юге Африки: Proteaceae, суккулентные Aizoaceae (аизовые: Lithops, Мesembryanthemum), а также центр распространения Ericaceae и Restionaceae (похожие на осоковые однодольные растения).
• Австралийское — царство, покрывающее в значительной степени Австралию: Myrtaceae (Eucalyptus, Leptospermum), Proteaceae (Banksiá), Casuarinaceae, Xanthorrho- eaceae(травяные деревья), а также центр разнообразия рода Acacia.
• Антарктическое — большей частью вымершее флористическое царство, но остатки его продолжают существовать на крайнем юге Америки, на южной оконечности Новой Зеландии и на субантарктических островах: буковым (Fagaceae) здесь соответствуют близкое семейство Nothofagaceae (Nothófagus), подушкообразная Azorella (Apiaceae); на антарктическом побережье в настоящее время произрастает только 2 коренных покрытосеменных растения: Deschampsia antárctica (Роасеае) и Colobanthus quitensis(Caryophyllaceae).
• Океаническое — флористическое царство Мирового океана и тихоокеанских островов с широко распространенными тропическими прибрежными родами и видами Cocos nucífera и Rhizophora sp. (мангры).
Эти флористические царства образуют основу для дальнейшего обсуждения флористических и растительных зон Земли (см. 15.2). В дальнейшем они могут делиться на упорядоченные иерархически флористические области, провинции, округа и районы (см. рис. 14.19, основные области распространения проникших в Центральную Европу геоэлементов)
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.