СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ IV. РАЗНООБРАЗИЕ
ГЛАВА 18. СЕМЕННЫЕ РАСТЕНИЯ
Покрытосеменные
Отдел покрытосеменных, или цветковых (Anthophyta), включает около 235 000 видов, т. е. намного больше, чем любая другая группа растений. Их вегетативная структура чрезвычайно разнообразна. Сюда относятся и гигантские эвкалипты, деревья высотой более 100 м со стволами почти20 м в обхвате (рис. 18-36), и мелкие ряски, просто устроенные плавающие однодольные, часто не превышающие в длину 1 мм (рис. 18-37). Некоторые покрытосеменные — лианы, высоко забирающиеся в полог дождевого тропического леса, другие — эпифиты, населяющие этот полог. Многие цветковые, например, кактусы, приспособлены к росту в крайне засушливых условиях. Уже более 100 млн. лет представители этого отдела господствуют на суше.
Рис. 18-25. Жизненный цикл сосны. Гаметофиты сильно редуцированы и в отношении питания зависят от спорофита. Незрелыми мужскими гаметофитами, состоящими из четырех клеток, являются пыльцевые зерна, которые переносятся ветром к женскому гаметофиту (мегагаметофиту), находящемуся в семязачатке. Не способные к активному движению спермии, образуемые прорастающими пыльцевыми зернами, доставляются к яйцеклеткам архегониев с помощью пыльцевых трубок (Вода как среда для их перемещения при этом не требуется). Семязачаток с мегагаметофитом внутри после оплодотворения созревает, становясь семенем. Характерный для сосен развитый подвесок ко времени полного развития зародыша дегенерирует. Семя сосны состоит из зародыша, семенной кожуры и мегагаметофита; представляющего собой запас питательных веществ

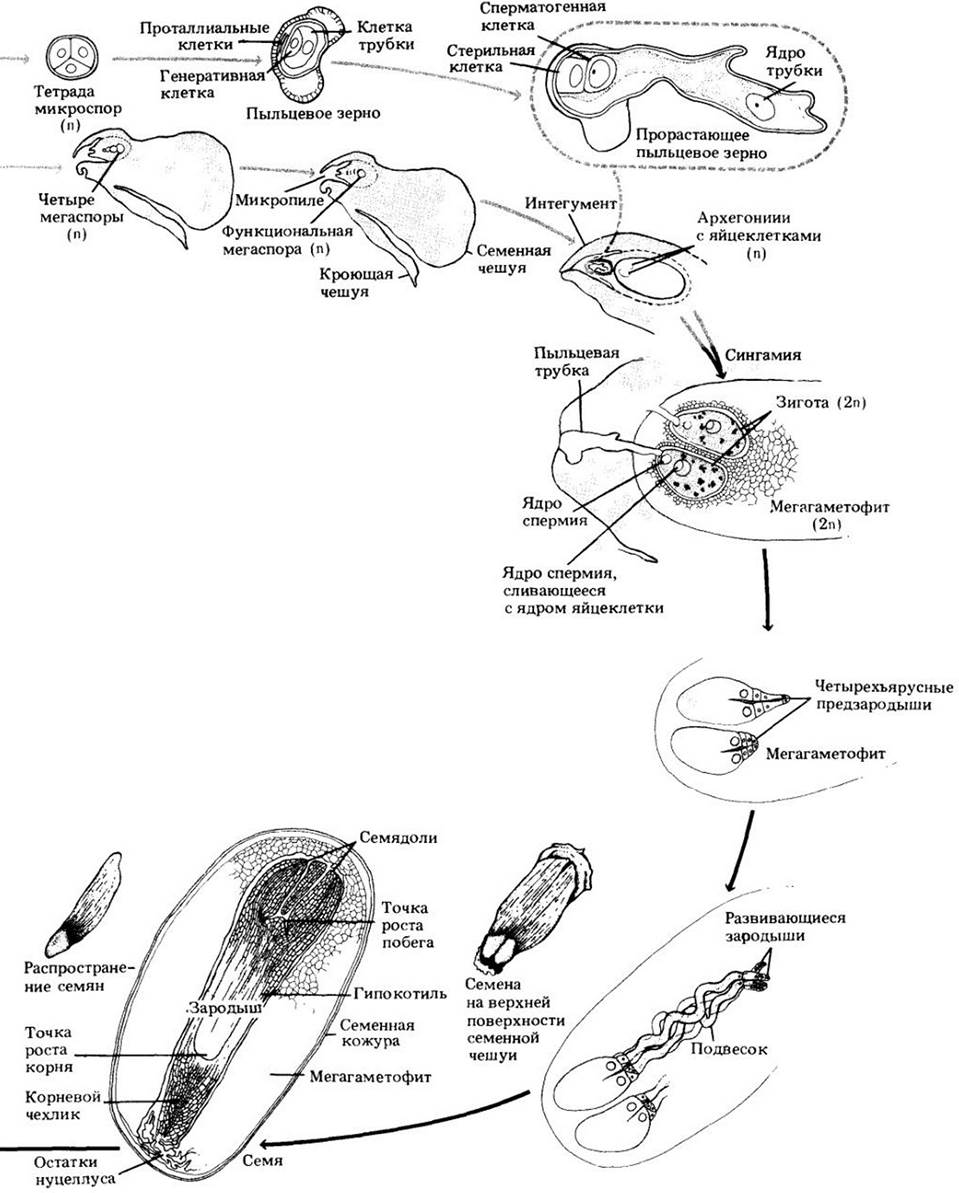
Рис. 18-26. Семена хвойных семейства тиссовых (Тахасеае) окружены мясистой чашей — ариллусом. Это образование привлекает птиц и других животных, которые, съедая его, распространяют семена. А. У тисса (Taxus), произрастающего в Северном полушарии, ариллусы красные. Б. Спорофиллы и микроспорангии микростробилов тисса. Мега- и микростробилы располагаются на разных деревьях. Ариллусы тисса ядовиты и являются наиболее частой причиной отравлений растениями у детей в США, хотя летальные исходы крайне редки
Рис. 18-27. Болотный кипарис (Taxodium distichum) — листопадный представитель семейства таксодиевых, растущий в болотах юго-востока США. У дерева, изображенного на этой, сделанной осенью фотографии, листья уже начали менять окраску. С ветвей у него в изобилии свисает так называемый бородатый мох, — в действительности цветковое растение Tillandsia usneoides, родственное ананасу

Рис. 18-28. Метасеквойя (Мetasequoia glyptostroboides). Этому дереву, растущему в провинции Хубэй в центральном Китае, более 400 лет

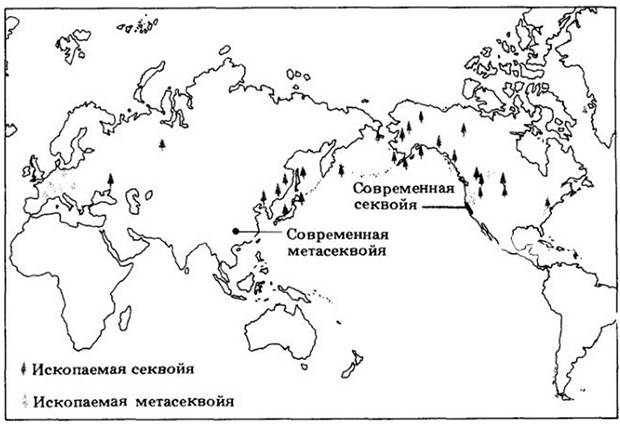
Рис. 18-29. Ископаемая веточка метасеквойи, которой около 50 млн. лет. На карте показано географическое распространение некоторых ныне живущих и ископаемых представителей семейства таксодиевых

Выделяют два класса покрытосеменных: Monocotyledones (однодольные) с примерно 65 000 видов (рис. 18-38) и Dicotylédones (двудольные) (рис. 18-39). Сходства между ними намного больше, чем различий, тем не менее это легко распознаваемые естественные группы. Среди однодольных такие хорошо известные растения, как злаки, лилии, ирисы, орхидеи, рогозы и пальмы. К двудольным относятся почти все знакомые нам деревья и кустарники (кроме хвойных) и многие травы. Основные различия между этими классами обобщены в табл. 18-1.
Таблица 18-1. Основные различия между однодольными и двудольными
Признаки |
Двудольные |
Однодольные |
Части цветка |
Обычно в мутовках по 4 или 5 |
Обычно в мутовках по три |
Пыльца |
В основе трикольпатная (с тремя бороздами или порами) |
В основе моноколь- патная (с одной бороздой или порой) |
Семядоли |
Две |
Одна |
Жилкование листьев |
Обычно сетчатое |
Обычно параллельное |
Первичные проводящие пучки в стебле |
Расположены кольцом |
Расположены сложным образом |
Настоящий вторичный рост (с участием камбия) |
Обычно имеется |
Отсутствует |
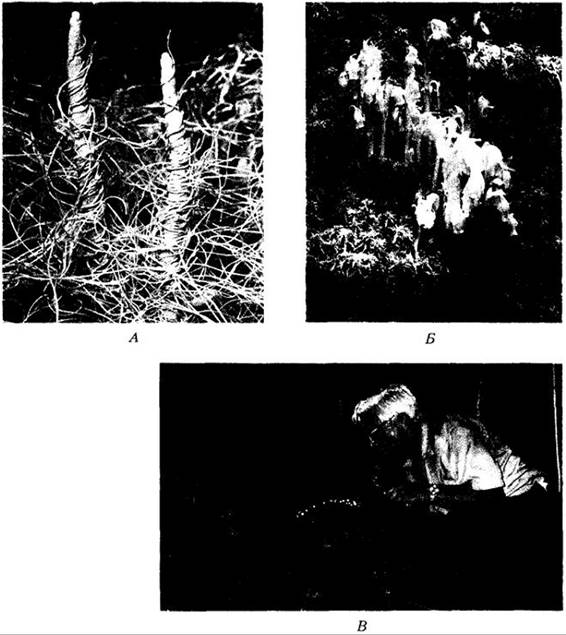
За немногими исключениями, покрытосеменные ведут независимый образ жизни, но среди них известны также паразиты и сапрофиты (рис. 18-40), в значительной степени или полностью лишенные хлорофилла. Недавно показано, что многие, если не все, цветковые «сапрофиты» облигатно связаны с микоризными грибами, которые одновременно взаимодействуют с зеленым фотосинтезирующим покрытосеменным. Гриб образует своего рода «мост», активно переносящий Углеводы от второго растения к бесцветному «сапрофиту». Существует около 2800 видов двудольных-паразитов, среди которых омела и повилика (рис. 18-40, A), и одно паразитное голосеменное (Parasitaxus из Новой Каледонии). Такие цветковые образуют особые поглощающие Органы, гаустории, которые проникают в Ткани хозяев.
Рис. 18-30. Женские и мужские растения замии карликовой (Zamia pumita) — единственного дикорастущего вида саговниковых в США. Ее целиком или отчасти подземные стебли вместе с главными корнями использовались в пищу индейцами семинолами. Две крупные серые «шишки» на переднем плане — женские, более мелкие коричневые — мужские

Рис. 18-31. А. Южноафриканский саговник Encephalartos altensteinii. Показан мужской экземпляр со стробилами. Б. Женский экземпляр Сусах siamensis. Вер хушка женской «шишки» удалена, чтобы показать семена на верхних поверхностях мегаспорофиллов

Рис. 18-32. А. Гинкго двулопастный. Б. Листья и мясистые семена гинкго на коротких побегах

Рис. 18-33. Крупные кожистые листья тропического гнетума напоминают листья некоторых двудольных. Виды этого рода — кустарники или деревянистые лианы в тропических и субтропических лесах. А. Собрания мегастробилов. Б. Собрания микростробилов и листья. В. Мясистые семена и листья. Фотографии Б и В сделаны в южновенесуэльском бассейне Амазонки

Рис. 18-34. Эфедра — единственный из трех родов гнетовых, встречающийся в США. А. Мужское растение Ephedra viridis (Калифорния), сильно разветвленный кустарник с чешуевидными, как у всех видов рода, листьями. Б. Микро- спорангиатные (мужские) стробилы Е. viridis. На стебле — чешуевидные листья. В. Мужские стробилы Ephedra trifurca (Аризона). Г. Женское растение Ephedra viridis с семенами
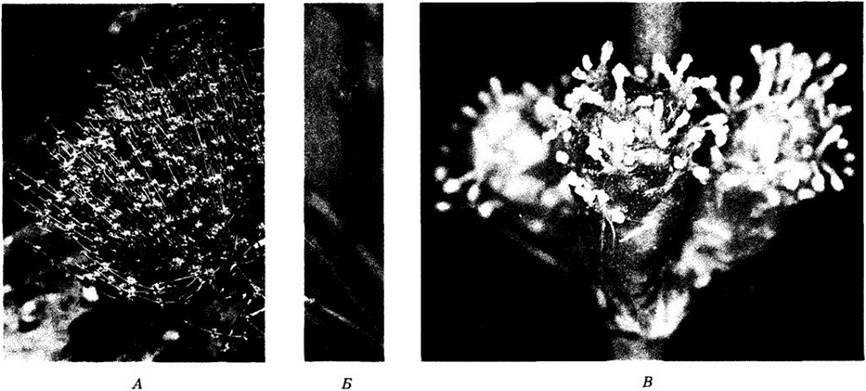
Рис. 18-35. Welwitschia mirabilis — представитель гнетовых, встречающийся только в пустыне Намиб и соседних областях юго-запада Африки. Вельвичия образует только два листа, не прекращающие роста в течение всей жизни растения. При этом они обламываются на верхушках и расщепяются продольно, поэтому кажется, что у старых растений листьев очень много. А. Крупное растение с семенами. Б. Микроспорангиатные стробилы. В. Стробилы с семязачатками; насекомое (клоп) высасывает сок из стробила. Вельвичия — двудомное растение


Рис. 18-36. Гигантский эвкалипт (Euca lyptus jacksonii) в Долине Гигантов на юго-западе Австралии. Огромные размеры этого покрытосеменного очевидны при сривнении с человеком, стоящим у его выжженного основания

Рис. 18-37. Рясковые (семейство Lemnaceae — самые мелкие цветковые растения. Черты их строения говорят о том, что это крайне редуцированные потомки ароидных (Аraceae)— семейства, в которое входят калла и филодендрон. А. Пчела, отдыхающая на плотном плавающем ковре, образованном тремя видами рясковых. Самое крупное растение здесь — Lemna gibba длиной 2 — 3 мм; поменьше — два вида Wolffia длиной до 1 мм. Б. Цветущее растение Wolffia borealis с круглым вогнутым рыльцем (похожим на крошечный бублик) и маленьким пыльником над ним, выступающими из центральной полости. Все растение меныие 1 м.ч длиной. В. Цветущее растение Lemna gibba; две тычинки и столбик выдаются из кармашка на верхней стороне растения

Рис. 18-38. Однодольные. А. Представитель семейства пальм кокос (Cocos nucífera) в Теуанатепеке (шт. Оахака, Мексика). Его плод (кокосовый орех) на самом деле костянка, а не орех (см. гл. 29). Б. Цветки и плоды банана (Musa х paradisiaca). Завязь у этого растения нижняя, на верхушке плода остается крупный рубец, оставленный опавшими частями цветка. В. Рис (Oryza sativa) из семейства злаков

Рис. 18-39. Двудольные. А. Кувшинка (Nymphaea odorata). Очень пахучий цветок этого вида с многочисленными тычинками и лепестками радиальносимметричный. Род Nymphaeaшироко распространен в тропических и умеренных областях по всему миру. Б. Кактус Carnegiea gigantea. Кактусы, которых насчитывается около 2000 видов, встречаются почти исключительно в Новом Свете. Их толстые мясистые стебли, запасающие воду, содержат Хлоропласты и фотосинтезируют вместо листьев. В. Печеночница (Hepática americana) расцветает в листопадных лесах ранней весной. Листочки околоцветника (их 6 —10) здесь не разделены на чашелистики и лепестки. Многочисленные тычинки и плодолистики расположены спирально
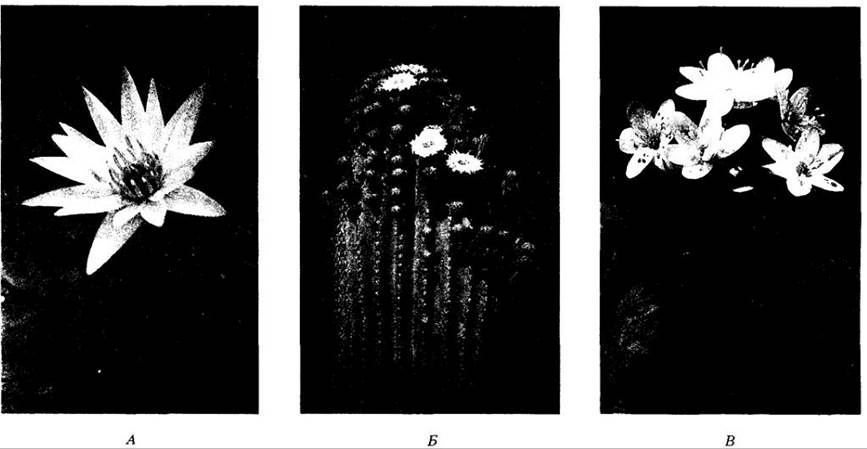
Рис. 18-40. Паразитические и сапрофитные покрытосеменные. Эти растения совсем или почти лишены хлорофилла и получают Питание за счет фотосинтеза других растений. А. Повилика (Cuscuta salina) — паразитическое ярко- оранжевое или желтое растение из семейства вьюнковых (Convolvulaceae). Б. Подъельник (Monotropa uniflora) — «сапрофит», получающий питательные вещества из корней других растений через гифы микоризного гриба. В. Самый крупный в мире цветок у вида Rafflesia arnoldii на горе Саго, Суматра. Растения этого рода паразитируют на корнях виноградовых (Vitaceae). Все растения на снимках — двудольные
В разделе V мы подробно рассмотрим структуру и развитие тела, т. е. спорофита цветковых растений, а остаток настоящей главы посвятим наиболее характерной особенности покрытосеменных — цветку — и процессу их размножения. В гл. 29 в деталях обсуждается эволюция этого отдела.
Цветок
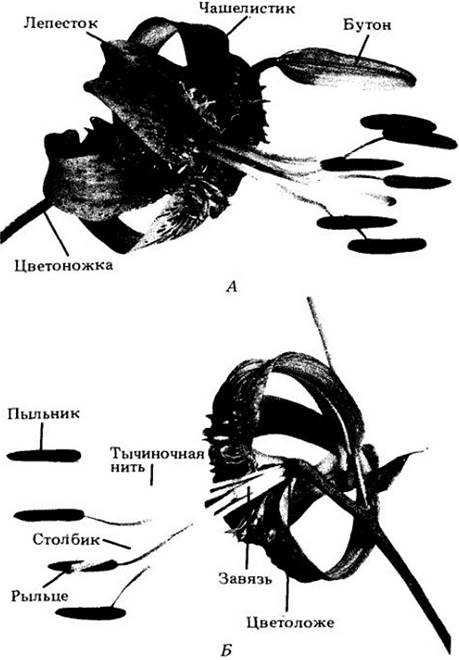
Цветок — это детерминированный ПОБЕГ, несущий спорофиллы (рис. 18-41). Название Angiospermae, переводимое на русский как «покрытосеменные», происходит от греческих слов angeion (сосуд) и sperma (семя), поскольку отличительная структура цветка — плодолистик — своего рода «сосуд», содержащий семязачатки (семяпочки), которые после оплодотворения развиваются в семена.
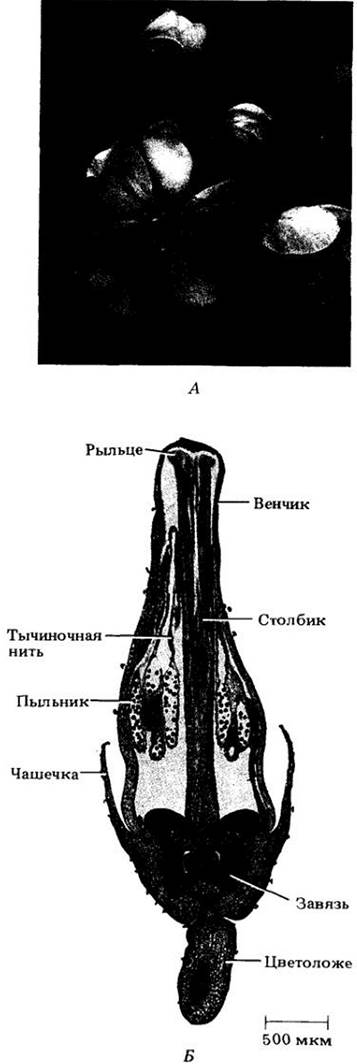
Рис. 18-41. Части цветка лилии (Lilium henryi). А. Ненарушенный цветок. Иногда, как и на рисунке, чашелистики и лепестки сходны друг с другом и называются вместе листочками околоцветника. Чашелистики прикреплены к цветоложу ниже лепестков. Б. Цветок, в котором удалены два листочка околоцветника и две тычинки, чтобы показать завязь. Гинецей состоит из завязи, столбика и рыльца, тычинка — из тычиночной нити и пыльника. Чашелистики, лепестки и тычинки здесь прикреплены к цветоложу ниже завязи; ее в этом случае называют верхней, а такой цветок — подпестичным (гипогинным)
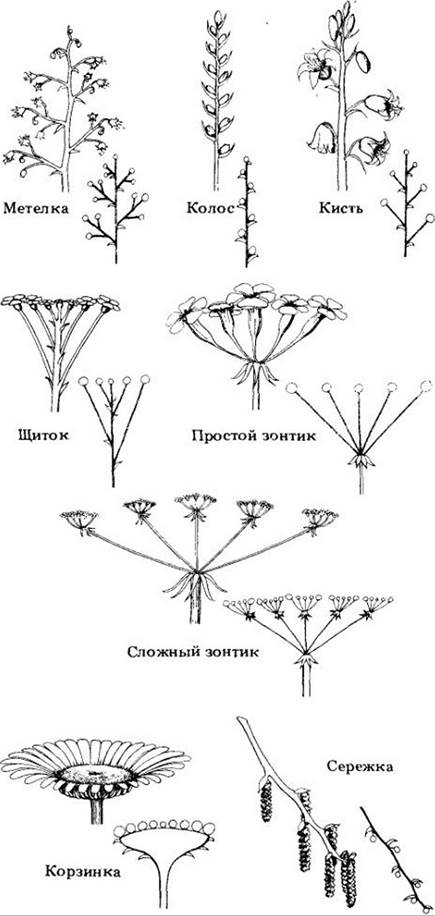
Цветки могут быть различным образом сгруппированы в соцветия (рис. 18-42 и 18-43). Ножка соцветия или одиночного цветка называется цветоносом, а ножка отдельного цветка в соцветии — цветоножкой. Цветоложе — часть цветоножки (цветоноса), к которой прикрепляются элементы (круги) цветка. Подобно любому концу побега, оно состоит из узлов и междоузлий, только междоузлия очень короткие, а узлы соответственно сильно сближены.
Рис. 18-42. Некоторые обычные типы соцветий покрытосеменных с упрощенными схемами
Многие цветки содержат два набора стерильных придатков, чашелистики и лепестки, прикрепленные к цветоложу ниже фертильных элементов, тычинок и плодолистиков (чашелистики отходят ниже лепестков, а тычинки — ниже плодолистиков). В совокупности первые образуют чашечку, а вторые — венчик, которые вместе составляют околоцветник. Чашелистики и лепестки по своей структуре во многом напоминают листья. Первые обычно зеленые, а вторые ярко окрашены, хотя у многих цветков оба круга сходны по цвету (рис. 18-41).
Тычинки, в совокупности, образующие андроцей (по-гречески «дом мужчины»), — это микроспорофиллы. У современных покрытосеменных, за немногими исключениями, они состоят из тонкого «стебелька» (тычиночной нити) с разделенными на две половинки пыльником, содержащим четыре микроспорангия, или пыльцевых мешка.
Плодолистики, или карпели, вместе составляющие гинецей (по-гречески «дом женщины»), — это мегаспорофиллы, сложенные в длину и заключающие внутри себя один или более семязачатков. В цветке может быть один или несколько карпелей, иногда полностью или частично сросшихся. Отдельный плодолистик или структуру, образованную сросшимися плодолистиками, часто называют пестиком из-за сходства формы с одноименным инструментом для растирания веществ в ступке.
У большинства цветков пестики дифференцированы на нижнюю часть — завязь, содержащую семязачатки, и верхнюю — рыльце, улавливающую пыльцу. Часто рыльце и завязь соединяются более или менее удлиненным образованием — столбиком. При срастании плодолистиков либо сливаются, либо сохраняются индивидуальные столбики или рыльца. Общая завязь сросшихся карпелей обычно (но не всегда) расчленена на два или более гнезд, в которых находятся семяпочки. Число гнезд, как правило, соответствует числу плодолистиков в гинецее.
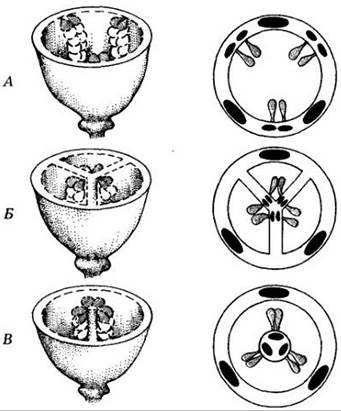
Места возникновения и прикрепления до зрелого состояния семязачатков в завязи называются плацентами. Их расположение (плацентация), а, следовательно, и размещение семяпочек у разных групп цветковых неодинаково (рис. 18-44). Оно бывает пристенным (париетальным), т. е семязачатки располагаются на стенке завязи или ее выпячиваниях. В других случаях семяпочки находятся на центральной колонке завязи, расчлененной на гнезда по числу плодолистиков (осевая плацентация). При свободной центральной плацентации семяпочки развиваются на центральной колонке, не связанной перегородками со стенкой завязи. И наконец, у некоторых цветков единственный семязачаток находится в самом основании одногнездной завязи (базальная плацентация). Эти различия имеют большое значение при классификации покрытосеменных.
Рис. 18-44. Типы плацентации. А. Париетальная. Б. Осевая. В. Свободная центральная. Базальная плацентация здесь не показана
Несмотря на существование общей основной структуры, между цветками разных растений существуют большие различия. Большинство их имеют и тычинки, и плодолистики; о таких цветках говорят, что они совершенные (обоеполые). Если либо тычинки, либо плодолистики отсутствуют, цветок называется несовершенным — соответственно пестичным или тычиночным (рис. 18-45). Если тычиночные и пестичные цветки находятся на одном растении (как, например, у кукурузы или дуба), вид называют однодомным, а если на разных (у ивы, падуба тусклого и др.) — двудомным. Другие семенные растения с органами, продуцирующими пыльцу и семена на разных особях (например, гинкго, саговники и можжевельники), также считаются двудомными.
Рис. 18-45. Тычиночные и пестичные цветки камнеплодника густоцветко- юго (Lithocarpus densiflora). Большин- meo представителей семейства буковых (Fagaceae), в которое входят и дубы Quercus), — однодомные растения; пычиночные и пестичные цветки у них Разделены, но расположены на одном и пом же дереве
Любой из кругов цветка — чашелистики, лепестки, тычинки или плодолистики — может у той или иной группы покрытосеменных отсутствовать. Если имеются все четыре круга, цветки называются полными, если отсутствует хотя бы один из них — неполными. Таким образом, несовершенный цветок является одновременно неполным, но не все неполные цветки несовершенны.
Части цветка располагаются либо спирально на более или менее удлиненном цветоложе, либо одинаковые элементы (например, лепестки) прикрепляются в узле мутовками. При этом внутри одной мутовки они могут срастаться между собой или прирастать к элементам других мутовок (например, тычинки часто прирастают к венчику). Если они не срастаются, для их описания употребляется приставка «раздельно-», в противоположном случае — «сростно-», или «спайно-» (соответственно в греческих терминах «апо-» и «сии-» или «сим-»). Например, раздельно- или сростнолистная чашечка, апокарпный или синкарпный гинецей.
Цветки различаются не только спиральным или мутовчатым расположением частей, но и уровнем их прикрепления к оси цветка по отношению к гинецею (рис. 18-46). Если прочие элементы отходят от цветоложа ниже его, говорят, что завязь верхняя, а цветок подпестичный (гипогинный) (см. рис. 18-41). У некоторых таких цветков чашелистики, лепестки и тычинки сливаются, образуя чашевидное продолжение цветоложа, называемое гипантием (околопестичные, или перигинные, цветки; рис. 18-47). При этом лепестки и тычинки кажутся отходящими от края чаши. В других случаях они как бы отрастают от верхушки так называемой нижней завязи (надпестичные, или эпигинные цветки; рис. 18-48).
Рис. 18-46. Типы цветков у трех обычных семейств двудольных, показывающие варианты положения завязи. У многих розовых (Rоsасеае) завязи верхние, а основания частей цветка срастаются в чашевидное расширение (гипантий). Такие цветки называют околопестичными (перигинными). У зонтичных (Ар1асеае) и кипрейных (Onagraceae) завязи нижние, т. е. части цветка прикреплены выше их. Цветки этих двух семейств называют надпестичными (эпигинными). Если гипантия нет, а части цветка прикрепляются ниже завязи, цветок называют подпестичным (гипогинным)
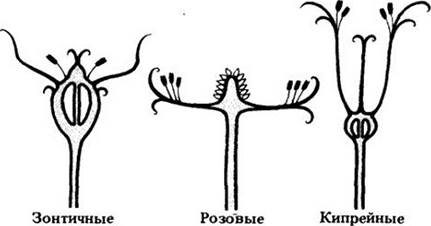
Рис. 18-47. А. Цветки вишни (Prunus) перигинные — их чашелистики, лепестки и тычинки прикрепляются к гипантию. На продольном разрезе бутона (Б) тычинки скучены в гипантии
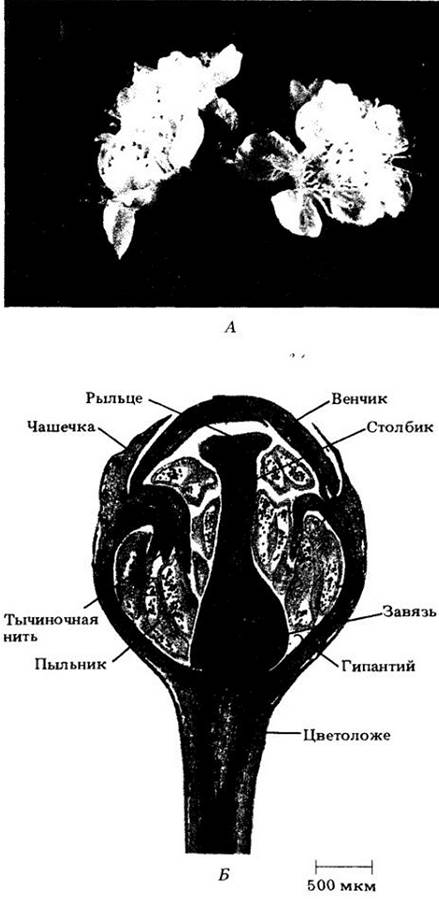
Рис. 18-48. А. Цветки яблони (Malus sylvestris) эпигинные — их чашелистики, лепестки и тычинки как бы отходят от верхушки завязи. Б. Разрез почти раскрытого цветка с еще не распрямленными тычинками
Наконец, говоря о разнообразии структуры цветков, следует упомянуть симметрию. В некоторых случаях венчик состоит из лепестков одинаковой формы, расходящихся на одинаковом расстоянии друг от друга от центра цветка, являющегося в результате радиально-симметричным. Такие цветки называют актиноморфными (от греческого корня actinos — «звезда»). В других случаях один или более элементов по крайней мере одной мутовки отличаются от других ее элементов и расположены двустороннесимметрично. Такие цветки называют зигоморфными (см. рис. 18-43, Б, В).
Жизненный цикл покрытосеменных
Гаметофиты цветковых очень сильно редуцированы в размерах — больше чем у любых других разноспоровых растений, включая голосеменные. Мужской в зрелом состоянии состоит только из трех клеток; а женский, погруженный в течение всей жизни в ткани спорофита, у большинства видов только из семи. Антеридии и архегонии отсутствуют. Опыление непрямое, т. е. пыльца откладывается на рыльце, а затем пыльцевая трубка доставляет два неспособных самостоятельно двигаться спермия к женскому гаметофиту. После оплодотворения семяпочка дает семя, заключенное в завязи, которая одновременно (иногда вместе с сопутствующими ей структурами) развивается в плод.
Микроспорогенез и микрогаметогенез
Микроспорогенез — образование микроспор в микроспорангиях, или пыльцевых мешках, пыльника. Микрогаметогенез — это развитие микроспоры в мужской гаметофит, или пыльцевое зерно.
На первых этапах дифференцировки пыльник состоит из однородной массы клеток и частично обособившейся эпидермы. Затем внутри него выделяются четыре группы фертильных, или спорогенных, клеток. Каждая такая группа окружена несколькими слоями стерильных клеток, которые превращаются в стенку пыльцевого мешка. В нее входят и питательные Клетки, которые снабжают питательными веществами развивающиеся микроспоры и образуют mane- тум, самый внутренний слой стенки (рис. 18-49). Спорогенные клетки становятся делящимися мейотически диплоидными микроспороцитами, дающими тетраду гаплоидных микроспор. Микроспорогенез завершается образованием одноклеточных микроспор.
Рис. 18-49. Два поперечных среза пыльников лилии (Lilium). А. Незрелый пыльник с четырьмя пыльцевыми мешками с микроспороцитами, окруженными mane- тумом. Б. Зрелый пыльник с пыльцевыми зернами. Перегородки между соседними пыльцевыми мешками разрушаются при его вскрытии

Во время мейоза образование клеточных стенок либо следует за каждым ядерным делением, либо разделяет протопласты четырех микроспор только после второго деления мейоза. Первый случай типичен для однодольных, второй — для двудольных. Следующим этапом является формирование основных признаков пыльцевых зерен (рис. 18-50). Они окружаются прочной наружной оболочкой — экзиной и целлюлозно-пектиновой внутренней оболочкой — интиной. Экзина состоит из очень стойкого вещества споро пол Ленина, по-видимому, образуемого частично тапетумом, а частично микроспорой. Интина откладывается протопластом микроспоры.
Рис. 18-50. Оболочка пыльцевого зерна служит для защиты мужского гаметофита во время его непростого путешествия от пыльника к рыльцу. Ее внешний слой, или экзина, состоит главным образом из спор о полле нина (полимера, состоящего в основном из каротиноидов). Жесткая и прочная экзина часто сложно скульптурирована. Скульптура оболочек вполне определенная и различная у разных видов, как видно на микрофотографиях, выполненных под сканирующим электронным микроскопом. А. Конский каштан (Aesculus hyppocastanum). Б. Лилия (Lilium longiflorum). В. Деталь поверхности пыльцевого зерна L. longiflorum. Г. Амброзия (Ambrosia psilosiachya). Пыльца амброзии является основной причиной аллергий в США. Шиповатые пыльцевые зерна, сходные с изображенным на рисунке, обычны в семействе сложноцветных (Asteraceae), к которому относится этот род
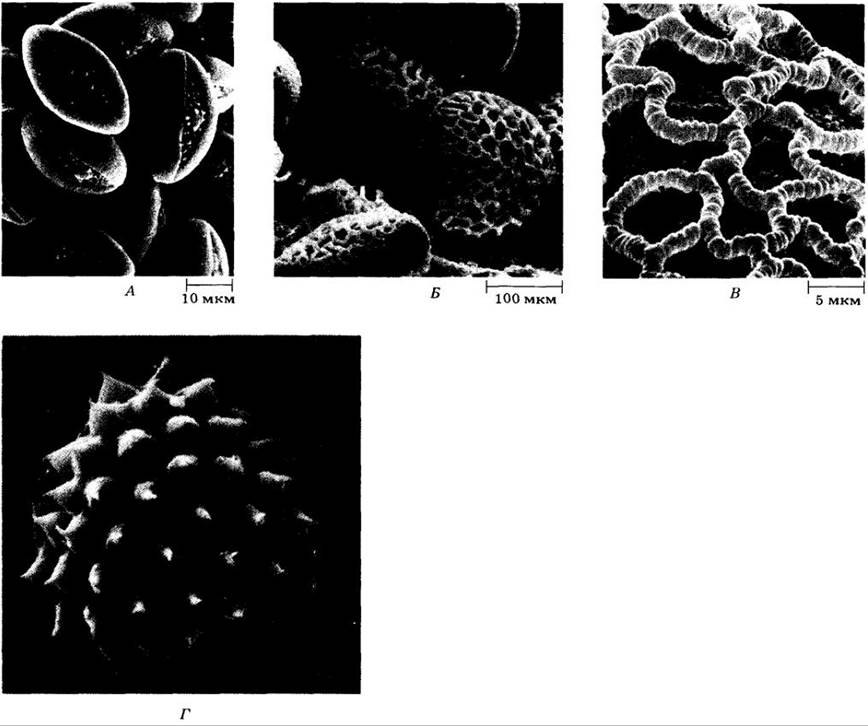
Размеры и форма пыльцевых зерен, как и у спор, сильно варьируют. Их диаметр составляет от менее 20 до более 250 нм. Они различаются также по числу и устройству пор, сквозь которые в конечном итоге прорастают пыльцевые трубки. Почти все семейства, многие роды и значительное число видов цветковых можно распознать по строению их пыльцевых зерен, в частности по их размеру, числу пор и скульптуре. В отличие от более крупных фрагментов растений — листьев, цветков и плодов — пыльцевые зерна прекрасно представлены в ископаемых отложениях благодаря химической природе их экзины. Таким образом, пыльца — ценный источник информации о господствовавших в прошлом растительности и климате.
И пыльцевые зерна, и Споры имеют спорополлениновую оболочку и являются продуктами мейоза. Однако в первых к моменту рассеивания в результате митоза образуются два или три ядра, а вторые содержат только одно. Споры прорастают через шов, проходящий по их центру, а пыльцевые зерна — через поры. Вследствие этого их часто нетрудно различить в ископаемом состоянии.
Микрогаметогенез у покрытосеменных единообразен и начинается с митотического деления одноядерной микроспоры, дающего две клетки внутри ее исходной оболочки. Одна из них называется клеткой трубки, а вторая — генеративной (рис. 18-51). Мужской гаметофит многих видов ко времени рассеивания пыльцы при вскрывании пыльника находится именно на этой двухклеточной стадии. У других видов генеративное Ядро делится, давая начало двум мужским гаметам, или спермиям, несколько раньше (рис. 18-52).
Рис. 18-51. Зрелое пыльцевое зерно лилии с двуклеточным мужским гаметофитом. Веретеновидная генеративная Клетка после митоза даст два спермин; более крупная клетка трубки образует позднее пыльцевую трубку
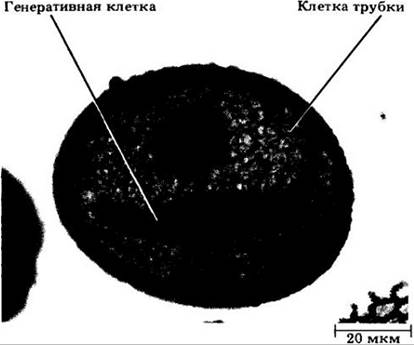
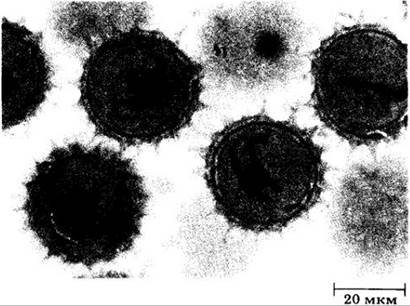
Рис. 18-52. Зрелые пыльцевые зерна с трехклеточными мужскими гаметофитами Silphium из сложноцветных. Перед опылением в каждом пыльцевом зерне находятся две нитевидные клетки спермиев, которые взвешены в цитоплазме более крупной клетки трубки. Другими словами, пыльца этого растения рассеивается на трехклеточной стадии, а не на двуклеточной, как у лилии с предыдущего рисунка
Мегаспорогенез и мегагаметогенез
Мегаспорогенез — это процесс формирования мегаспоры в нуцеллусе (мегаспорангии). Мегагаметогенез — это развитие мегаспоры в женский гаметофит.
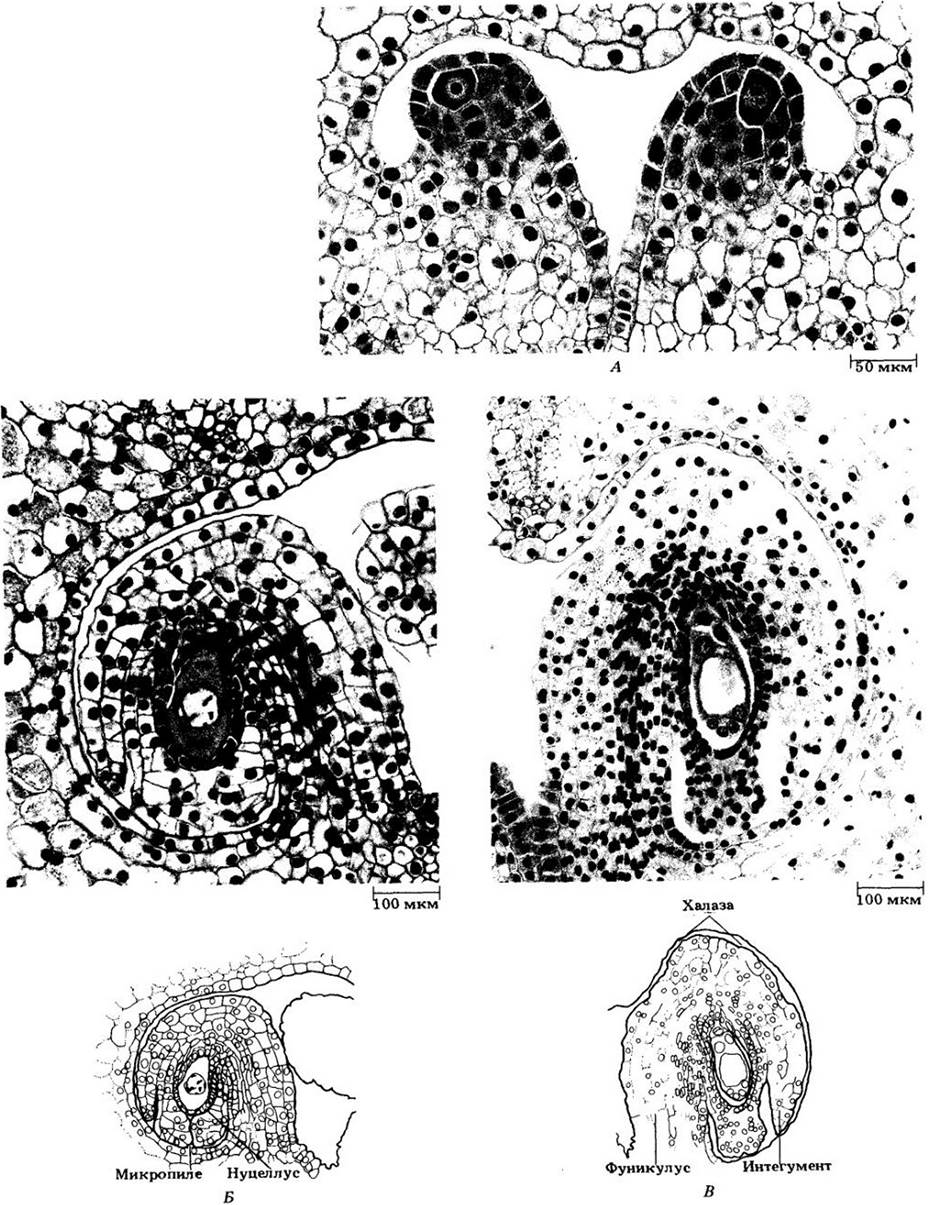
Семяпочка — относительно сложное образование, состоящие из семяножки (фуникулуса), несущей нуцеллус, заключенный в один или два интегумента. В зависимости от вида на плацентах развиваются от одного до многих семязачатков. Развивающаяся семяпочка первоначально состоит целиком из нуцеллуса, но вскоре возникают один или два покровных слоя (интегумента) с небольшим отверстием, микропиле, на одном конце (рис. 18-53).
На ранней стадии развития семязачатка в нуцеллусе возникает единственный диплоидный мегаспороцит. Он делится мейотически (рис. 18-53,15), давая четыре гаплоидные мегаспоры, обычно расположенные линейной тетрадой. Этим завершается мегаспорогенез. Три мегаспоры обычно разрушаются, а четвертая, наиболее удаленная от микропиле, развивается в женский гаметофит.
Функциональная мегаспора вскорее начинает увеличиваться за счет нуцеллуса, а ее ядро трижды делится митотически. В конце третьего митоза восемь дочерних ядер располагаются по четыре двумя группами — вблизи микропиляр- ного конца мегагаметофита, а также на противоположном, халазалъном, конце. По одному ядру из каждой группы мигрирует к центру восьмиядерной клетки; они называются полярными. Три оставшихся у микропилярного конца ядра образуют яйцевой аппарат, состоящий из яйцеклетки и двух клеток-синергид. У халазального конца также идет формирование вокруг расположенных здесь ядер клеточных оболочек, и возникают так нызываемые клетки-антиподы. Полярные ядра остаются в двуядерной центральной клетке. Такая восьмиядерная семиклеточная структура и является зрелым женским гаметофитом, называемым зародышевым мешком (рис. 18-53,19).
Рис. 18-53. Лилия. Некоторые стадии развития семяпочки и зародышевого мешка. А. Две семяпочки на ранней стадии, каждая с одним крупным мегаспороцитом. Интегументы еще не начали развиваться. Б. Семяпочка уже с интегументами. Мегаспороцит в первой профазе мейоза. В. Семяпочка с восьмиядерным зародышевым мешком (здесь видны только шесть ядер). Полярные ядра еще не мигрировали к его центру
Описанный выше тип его развития наиболее распространен, однако примерно у трети исследованных с этой точки зрения видов покрытосеменных он иной.
Опыление и Оплодотворение
При вскрывании (рассеивании содержимого) пыльников пыльцевые зерна переносятся на рыльца многими способами (см. гл. 29); сам процесс этого переноса называется опылением. Вступив в контакт с рыльцем, пыльцевые зерна поглощают дополнительное количество воды из клеток его поверхности (по градиенту водного потенциала). После насыщения водой они прорастают в пыльцевую трубку. Генеративная клетка, если она еще не поделилась, вскоре делится, образуя два спермия. Проросшее пыльцевое зерно с ядром трубки и двумя спермиями и соответствует зрелому мужскому гаметофиту (рис. 18-55).
Рыльце и столбик структурно-физиологически устроены так, чтобы облегчить прорастание пыльцевого зерна и рост пыльцевой трубки. Поверхность многих рылец образована главным образом железистой тканью (рылъцевая ткань), выделяющей сахаристый раствор. Эта ткань связана с семяпочкой проводниковой (трансмиссионной) тканью, своего рода дорожкой сквозь столбик для растущих пыльцевых трубок. В некоторых столбиках проводниковой тканью выстланы изнутри открытые канальцы и пыльцевые трубки растут либо поверх этой выстилки, либо среди ее клеток. Однако у большинства покрытосеменных столбики выпол-
ненные (сплошные) с одним или несколькими тяжами проводниковой ткани между рыльцем и семяпочками. Пыльцевые трубки здесь в зависимости от вида растения растут либо среди ее клеток, либо внутри их толстых стенок.
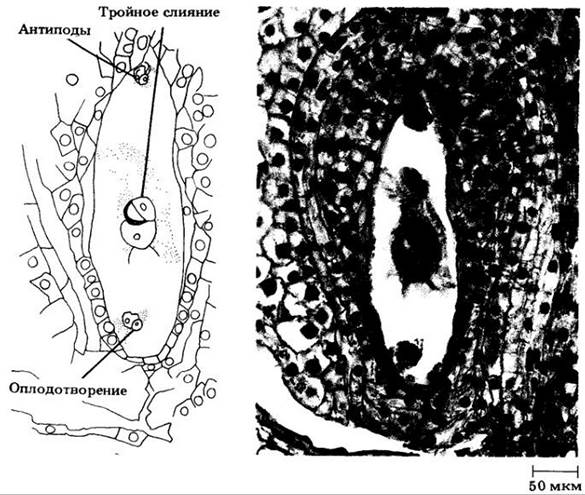
Обычно пыльцевая трубка входит в семяпочку через микропиле и проникает в одну из синергид, которая начинает отмирать вскоре после опыления, но раньше, чем пыльцевая трубка достигает зародышевого мешка. Два спермия и ядро трубки выпускаются в эту синергиду через открывающуюся в пыльцевой трубке субтерминальную пору. Затем ядро одного из спермиев проникает в яйцеклетку, а другого — в центральную клетку, где сливается с двумя полярными ядрами (рис. 18-56). В этом отличие от голосеменных, у которых функционален только один из двух спермиев, сливающийся с яйцеклеткой, в то время как второй дегенерирует. Слияние одного спермия с яйцеклеткой, а другого с полярными ядрами — двойное оплодотворение — и представляет собой уникальную особенность покрытосеменных. Как отмечалось выше, «настоящее» оплодотворение, или сингамия, подразумевает только слияние гамет, в данном случае яйцеклетки и спермия, в результате чего образуется диплоидная зигота. Соединение другого спермия с двумя полярными ядрами, т. е. тройное слияние, дает триплоидное первичное ядро эндосперма. Ядро клетки трубки тем временем отмирает, оставшаяся синергида и антиподы отмирают вместе с ним или на ранних стадиях дифференцировки зародышевого мешка.
Развитие семени и плода
После двойного оплодотворения начинается несколько процессов: первичное ядро эндосперма делится, образуя эндосперм; зигота развивается в зародыш; интегументы превращаются в семенную кожуру, а стенка завязи и связанные с ней структуры формируют плод.
В противоположность эмбриогенезу большинства голосеменных, начинающемуся со свободноядерной стадии, у покрытосеменных, этот процесс протекает, скорее, как у споровых сосудистых растений, т. е. первое ядерное деление зиготы сопровождается образованием клеточной оболочки. На ранних стадиях развития последовательность клеточных делений у зародышей двудольных и однодольных сходна; в обоих случаях формируются сферические тела. Позже появляется различие: зародыш двудольных развивает две семядоли, а однодольных — только одну. Подробности эмбриогенеза покрытосеменных рассмотрены в разделе V.
Образование эндосперма начинается с митотического деления его первичного ядра, происходящего обычно раньше первого деления зиготы. При этом у некоторых покрытосеменных, образованию клеточных оболочек предшествует различное количество делений свободных ядер (нуклеарный тип образования эндосперма); у других и первый, и последующие митозы сопровождаются цитокинезом (целлюлярный тип образования эндосперма). Хотя формирование эндосперма происходит разными путями, функция возникающей ткани всегда одна: обеспечение развивающегося зародыша, а во многих случаях и молодого проростка необходимыми питательными веществами. В семенах некоторых групп покрытосеменных нуцеллус разрастается в запасную ткань, называемую периспермом. Некоторые семена содержат и эндосперм, и перисперм, например, у свеклы (Beta). Однако у многих двудольных и некоторых однодольных вся запасная ткань или большая ее часть поглощается развивающимся зародышем еще до того, как семя впадает в состояние покоя (у гороха, бобов и др.). Зародыши в таких семенах обычно развивают заполненные питательными веществами мясистые семядоли. Основные питательные вещества, запасаемые в семенах, — углеводы, Белки и ЛИПИДЫ.
Семена голосеменных и покрытосеменных различаются происхождением этих веществ. У первых их дает женский гаметофит, у вторых — по крайней мере в начале — эндосперм, не являющийся ни гаметофитной, ни спорофитной тканью.
Развитие семяпочки в семя сопровождается превращением завязи (а иногда и других частей цветка или соцветия) в плод. В ходе этого стенка завязи (околоплодник, или перикарпий) часто утолщается и дифференцируется на отдельные слои — внешний экзокарпий (внеплодник), средний мезокарпий (межплодник) и внутренний эндокарпий (внутриплодник) (иногда только на экзо- и эндокарпий), — обычно лучше заметные в мясистых, чем в сухих, плодах. Плоды более подробно обсуждаются в гл. 29.
Жизненный цикл покрытосеменных представлен на рис. 18-54.
Рис. 18-54. Жизненный цикл сои (Glycine mах) — представителя цветковых. Из семени после его прорастания развивается взрослый спорофит, который, как и у других покрытосеменных, в конечном итоге образует цветки. В пыльниках цветка формируются материнские клетки микроспоры, или микроспороциты. Мейоз каждой из них дает начало четырем гаплоидным микроспорам, которые делятся один раз, образуя клетку трубки и генеративную клетку. Такая двуклеточная структура представляет собой незрелый микрогаметофит, или пыльцевое зерно. Во время его прорастания или еще раньше генеративная клетка делится на два спермин. Они доставляются к яйцевому аппарату пыльцевой трубкой. Проросшее пыльцевое зерно с ядром трубки и двумя спермиями — это зрелый мужской гаметофит. В семяпочке развивается и в конечном итоге делится мейотически материнская клетка мегаспоры, или мегаспороцит, давая начало четырем мегаспорам, три из которых отмирают. Четвертая развивается в женский гаметофит, в зрелом состоянии являющийся семиклеточным восьмиядерным зародышевым мешком. Пыльцевое зерно прорастает на рыльце, давая пыльцевую трубку, растущую вниз к завязи через столбик и входящую в семяпочку через микропиле. Ядро одного спермия из пыльцевой трубки сливается с яйцеклеткой в зиготу, а ядро второго — с двумя полярными ядрами зародышевого мешка в триплоидное первичное ядро эндосперма. Такое двойное оплодотворение известно только у покрытосеменных. В зародышевом мешке развивается зародыш, интегументы семяпочки превращаются в семенную кожуру. В конце концов семена рассеиваются


Рис. 18-55. Зрелый мужской гаметофит купены (Polygonatum). В пыльцевой трубке видны спермии и ядро трубки
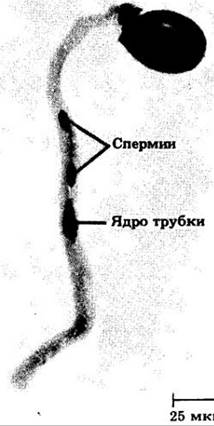
Рис. 18-56. Двойное оплодотворение у лилии. Слияние ядер спермин и яйцеклетки — «настоящее» оплодотворение — видно в нижней части микрофотографии. Выше — тройное слияние ядра другого спермия и двух полярных ядер
Последнее обновление: 05/02/2024
Редакционная и учебная адаптация: Данный материал сведен на основе первоисточника/оригинального текста. Команда проекта осуществила редакционную обзорную обработку, исправление технических неточностей, структурирование разделов и адаптацию содержания к учебному формату.
Что было обработано:
- устранение форматных дефектов (OCR-ошибки, разрывы структуры, дефектные символы);
- редакционное упорядочивание содержания;
- унификация терминов в соответствии с академическими источниками;
- проверка соответствия фактических утверждений текста первоисточнику.
Все упоминания об авторе, годе издания и происхождении первичного текста сохранены в соответствии с источником.